| 【緒言】 哺乳動物では、胎生期から閉経に至るまで絶え間なく卵胞発育が続けられる。しかし、99.9%の卵胞は発育の様々な段階で卵胞閉鎖(atresia)と呼ばれる変性に至る。胎生初期には扁平な一層の上皮細胞に覆われた原始卵胞と呼ばれる状態で、卵は減数分裂のdiplotene stageで止まっているが、胎生20週頃より卵胞発育が開始する。卵細胞の増大とともに卵を取り囲んでいた顆粒膜細胞が円柱状になり(一次卵胞)、顆粒膜細胞層が多層化(3-5層:二次卵胞、6層以上:三次卵胞)する。さらに顆粒膜細胞層の間隙に形成された卵胞腔が増大して排卵に至る。卵胞閉鎖は、形態学的には卵胞顆粒膜細胞の核濃縮から始まり、進行すると顆粒膜細胞層の剥離と変性、基底膜の断片化に至る。卵胞閉鎖は発育のあらゆる段階で認められるが、特に二次卵胞から三次卵胞に至る時期に大部分が閉鎖に至るとされている。これまで閉鎖卵胞(atretic follicle)の生化学的特性については、DNA合成の減少、エストロゲン産生の減少や、下垂体ホルモン結合能の減少など様々な報告があるが、閉鎖に至る卵胞が選択される機序については未だ明らかでない。最近、閉鎖卵胞において顆粒膜細胞のアポトーシスが起きていること、顆粒膜細胞と黄体細胞はアポトーシスの指標の一つであるDNA断片化を起こすのに必要なCa2+,Mg2+依存性endonuclease活性を持ち、その活性は卵巣ステロイドホルモンやIGF-I,EGFなどのgrowth factorにより調節されることがなどが報告されている。 Fas/APO-1/CD95(以下Fas)はTNF(tumor necrosis factor)/NGF(nerve growth factor)受容体ファミリーに属する45-kDaの膜貫通型糖蛋白質であり免疫系細胞のみならず、心臓、肝臓、卵巣などにも多量に発現していることがmRNAレベルで報告されている。これに対するligandであるFas ligandは31-kDaの膜貫通型蛋白質であり、TNF/NGF受容体ファミリーのそれぞれのligandとhomologyが高い。Fas-Fas ligand間の相互作用によりアポトーシスが誘導されることが免疫系細胞では報告されてきた。卵巣より分離した顆粒膜細胞にFasが多量に発現していることから、卵胞閉鎖への関与が示唆されたが、その卵巣内における局在や役割については明らかでない。 本研究では、FasおよびFas ligandの蛋白質および遺伝子レベルでの発現と局在をin vivoで明らかにした。さらに、Fas-Fas ligand間の相互作用が顆粒膜細胞のアポトーシスを誘導することをin vitroで示した。 【方法と結果】 Fas-Fas ligandシステムの卵巣内における局在を明らかにするために、FasおよびFas ligandに対する特異抗体を作成した。マウスFasの細胞外領域および細胞内領域それぞれに相当する2種類の合成ペプチド(P2およびP4)を作成し、これをウサギに投与しポリクローナル抗体を得た。これらの抗Fas抗体(anti-P2とanti-P4)を用いたウエスタンプロット法により、pregnant mare serum gonadotropin(PMSG)を投与した幼若雌ラットや成熟雌ラットの卵巣より抽出した蛋白質中に、45kDaのFasに相当する分子量のバンドが検出された。一方、ラットFas ligandの細胞内領域に相当する合成ペプチド(P5)を作成し得られた抗Fas ligand抗体(anti-P5)を用いて、31kDaのFas ligandに相当する分子量のバンドが卵巣全体より抽出した蛋白質および卵より抽出した蛋白質中に検出されたが、顆粒膜細胞を分離して抽出した蛋白質中には特異的なバンドは認められなかった。以上よりFasおよびFas ligandの卵巣における蛋白質レベルでの存在が確認された。 次に、FasおよびFas ligandの卵巣内おける局在と卵胞閉鎖との関連を検討するため、PMSGを投与した幼若ラットの卵巣切片を作成した。PMSGを投与した幼若ラットの卵巣は、様々な発育段階の健康な発育卵胞および閉鎖卵胞が多数観察されるため卵胞発育の解析によく用いられるモデルである。その切片上でヘマトキシリン・エオジン染色を施行し卵胞の形態を観察したところ、一次卵胞および一部の二次卵胞では顆粒膜細胞の核濃縮より先に卵の変性が生じていたが、その他の二次卵胞、三次卵胞では顆粒膜細胞の核濃縮が先行していた。原始卵胞は形態的には均一で、卵胞閉鎖に至るものを判別することはできなかった。 ヘマトキリリン・エオジン染色で卵胞の状態を判定した切片と連続する切片上で、アポトーシスの指標の一つであるDNA断片化を検出するために、terminal deoxy-UTP nick end labeling(TUNEL)を施行して顆粒膜細胞のアポトーシスの有無を検討した。顆粒膜細胞の核濃縮が認められる閉鎖卵胞では、顆粒膜細胞の核がTUNELにより染色され、アポトーシスを起こしていることが示唆された。健康な発育卵胞と閉鎖卵胞のいずれにおいても卵はTUNELにより染色されなかった。原始卵胞でTUNELにより染色されるものはなかった。 さらに、連続切片上で抗Fas抗体(anti-P4)を用いた免疫組織化学を施行した。原始卵胞で抗Fas抗体に反応するものはなかった。顆粒膜細胞の核濃縮により卵報閉鎖と判断される二次卵胞および三次卵胞において顆粒膜細胞が特異的に染色されたが、健康な発育卵胞は染色されなかった。免疫染色の特異性はP4ペプチドを用いた吸収テストで確認された。顆粒膜細胞層の剥離を伴った卵胞閉鎖が進行した卵胞では非特異的抗体結合が強く、Fasの発現を組織学的に証明することはできなかった。卵にも抗Fas抗体の結合が認められたが、吸収テストで特異的染色であることが示されなかった。抗Fas抗体で染色された卵胞の90%以上はTUNELも陽性であった。また、抗Fas抗体陰性の卵胞はすべてTUNEL陰性であった。anti-P2抗体を用いた免疫染色でも同様の結果が得られた。mRNAレベルでのFasの発現を検討するため、digoxigeninで標識したFas cDNA probeを用いてnon-radioactive in situ hybridizationを施行した。Fas mRNAは閉鎖卵胞の顆粒膜細胞に発現していたが、卵には検出されなかった。 抗Fas ligandを用いた免疫染色では、正常・閉鎖の区別無く一次から三次のすべての卵胞の卵表面が三日月状に特異的に染色された。原始卵胞では染色は認められなかった。Fas ligand cDNA probeを用いたnon-radioactive in situ hybridizationでは卵巣切片上でFas ligand mRNAは検出されなかった。 PMSGを投与した幼若ラットから分離した顆粒膜細胞の培養系でFasの免疫染色を施行したところ、これまでの報告と同様にIFN 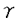 の存在下でFasの発現が促進されることが示された。 の存在下でFasの発現が促進されることが示された。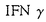 で前培養しFasを発現した培養顆粒膜細胞を透明帯除去卵と共培養すると、顆粒膜細胞のアポトーシスの頻度が増加した。培養顆粒膜細胞のアポトーシスは、Hoechst33342により染色した核の断片化を指標とした。この際、同時に抗Fas抗体(anti-P2抗体)を添加するとアポトーシス誘導が抑制され、抗Fas抗体がFas-Fas ligand間の相互作用を阻害していると考えられた。 で前培養しFasを発現した培養顆粒膜細胞を透明帯除去卵と共培養すると、顆粒膜細胞のアポトーシスの頻度が増加した。培養顆粒膜細胞のアポトーシスは、Hoechst33342により染色した核の断片化を指標とした。この際、同時に抗Fas抗体(anti-P2抗体)を添加するとアポトーシス誘導が抑制され、抗Fas抗体がFas-Fas ligand間の相互作用を阻害していると考えられた。 【考察】 Fasは閉鎖卵胞の顆粒膜細胞に発現しており、顆粒膜細胞のアポトーシスと密接に関連していた。一方、Fas ligandは、一次から三次のすべての卵胞の卵表面に三日月状に発現が認められ、卵の極性による差が示唆された。Fas ligandは遺伝子レベルでは検出されなかったが、このmRNAがin situ hybridizationの感度以下である可能性と、卵のより未熟な段階で産生され蓄積されている可能性が考えられた。一次卵胞や、一部の二次卵胞では、卵の変性が顆粒膜細胞のアポトーシスに先行しており、これらの卵胞ではFasの発現は認められなかった。未熟な発育段階の卵胞閉鎖では、Fas-Fas ligandシステムとは異なる機序が働いていると考えられた。 培養顆粒膜細胞においてがFas発現を増加させることは以前にも報告があったが、同様の方法でFasを発現させた顆粒膜細胞と透明体除去卵との共培養により顆粒膜細胞のアポトーシスを誘導することができた。Fas ligandは免疫組織学的に卵表面にのみ検出されたが、Fas ligandとhomologyが高いTNFで報告されているように、膜表面型以外に分泌型がある可能性も考えられる。また、卵巣には免疫系細胞の浸潤が頻繁に認められるため、それらがFas ligandを供給しているかもしれない。 【結論】 卵巣顆粒膜細胞膜表面上のFasと卵に発現するFas ligandとの相互作用が顆粒膜細胞のアポトーシスを誘導し、卵胞閉鎖に至る可能性が示された。顆粒膜細胞のFas発現の調節機構は明らかでないが、などのサイトカインの関与が示唆された。 多くの卵胞が閉鎖に至る中で、発育し排卵に至る卵胞がどのような機序で選択されるのかは未だ明らかでない。その解明は、臨床的にも卵胞発育の異常がみられる多嚢胞卵巣症侯群や早発閉経などの疾患の病態解明や不妊症に対する治療の発展に貢献するものと考えられる。 |