| 内容要旨 | | 研究目的 ランゲルハンス細胞(LC)は、骨髄由来の抗原提示細胞で表皮に定住する。LCは外界から抗原を取り込んだ後に所属リンパ節へ移動し、T細胞に外来抗原を提示しT細胞を活性化する。このT細胞の活性化はMHC class II-T細胞レセプターを介する系、LCのB7-1、B7-2、T細胞のCD28、CTLA-4を介する系によって調節されることが明らかとなっているが、さらに近年LCのCD40とT細胞のCD40リガンドとの関係が報告されている。従ってこれらの分子の調節作用を検討することは、LCの抗原提示能の解析に重要と思われる。またGM-CSF,TNF-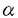 などのサイトカインはLC活性化を誘導する。いっぼうこれら分子への調節機序の検討は、1-2%のLCを含んだ表皮細胞(EC)の細胞成分を用いたものが多い。従ってECの大部分を占め多数のサイトカインを産生するケラチノサイト(KC)による影響は否定できない。今回、新鮮LGを高純度(95%)で精製し、新鮮LCと培養LCにおけるB7-1,B7-2、CD40分子の発現を比較検討した。さらにこの系にサイトカインおよび種々の免疫抑制剤を加えて、培養LCのこれらの分子の発現作用について解析を加えた。またLCのサイトカインに対するレセプターの発現についても検討した。さらにステロイドによって誘導されるB7-1,B7-2,CD40の発現阻害に及ぼすサイトカインの影響についても検討した。 などのサイトカインはLC活性化を誘導する。いっぼうこれら分子への調節機序の検討は、1-2%のLCを含んだ表皮細胞(EC)の細胞成分を用いたものが多い。従ってECの大部分を占め多数のサイトカインを産生するケラチノサイト(KC)による影響は否定できない。今回、新鮮LGを高純度(95%)で精製し、新鮮LCと培養LCにおけるB7-1,B7-2、CD40分子の発現を比較検討した。さらにこの系にサイトカインおよび種々の免疫抑制剤を加えて、培養LCのこれらの分子の発現作用について解析を加えた。またLCのサイトカインに対するレセプターの発現についても検討した。さらにステロイドによって誘導されるB7-1,B7-2,CD40の発現阻害に及ぼすサイトカインの影響についても検討した。 方法 実験1 Balb/cマウスの躯幹の皮膚を採取し、dispase、deoxyribonuclease処理にて表皮細胞(EC)浮遊液を作成した。このEC浮遊液を抗Ia抗体で処理し、あらかじめ抗mouse抗体をコートしたシャーレ上で反応させ、シャーレ上の接着細胞を採取し精製LCとした(>95%)。このようにして採取した新鮮LC(fLC)をFACSを用いてそのIa,NLDC145,CD14などの発現を培養(36時間)前後で比較した。また形態的にも検討した。 実験2 上記にて採取したLCを用いてmRNAを採取し、cDNAを合成し、サイトカインレセプターに対するprimerを用いてRT-PCRを行い、LCのサイトカインレセプター(M-CSFR,TNF-R,GM-CSFR)を検討した。またLCのサイトカイン発現についても検討を加えた。このとき表皮細胞浮遊液よりLC,Thy-1表皮陽性樹状細胞(DETC)の順にenrichementを行い、残った分画をKCとして用いた。 実験3 精製LC(1X105/well)を、サイトカイン(M-CSF,GM-CSF,TNF-)を加えて36時間培養し、培養LC(cLC)上のB7-1,B7-2,CD40の発現の変化をFACSを用いて検討した。サイトカインによるLCのB7-1,B7-2,CD40発現の調節作用を以下のように解析した。 %modulation of MFI(mean fluorescence intensity)=(MFI of LC treated with cytokine or immunosuppressants/MFI of LC trated with medium only)x100% 実験4 精製LCを用いて種々の免疫抑制剤(dexamethasone(DEX),cyclosporinA(CsA),FK506)を加えて、36時間培養し、cLC上のB7-1,B7-2,CD40分子の発現の変化をFACSを用いて解析した。 実験5 精製LCを用いてDEXとサイトカイン(M-CSF,GM-CSF,TNF-)を加えて36時間培養し、cLC上のB7-1,B7-2,CD40の発現をFACSを用いて検討した。 結果 今回パンニング法を用いてMHC classII(Ia)抗原陽性細胞が95%以上を占めるLCを採取し、この精製LCのB7-1,B7-2,CD40の発現に対するサイトカイン、免疫抑制剤の作用を解析した。 実験1 EC由来のIa陽性細胞は95%以上のpurityで採取され、これを各実験に用いた。fLC,cLCのNLDC145,CD14,CD11a,CD11b,CD11cの発現はfLCではNLDC145-,CD14-,CD11a+,CD11b+,CD11c+,cLCではNLDC145+,CD14-,CD11a+,CD11b+,CD11c+であり、pureなLCと考えた。 実験2 fLCのサイトカインレセプターの発現をRT-PCRで検討した。fLCにM-CSFR,GM-CSFR,TNF-RII mRNAの発現を認めた。またIL-1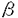 ,TNF- mRNAなどのサイトカインmRNAの産生がfLCで確認された。しかしIL-1,GM-CSF,M-CSFmRNAなどは確認されなかった。一方DETCでは ,TNF- mRNAなどのサイトカインmRNAの産生がfLCで確認された。しかしIL-1,GM-CSF,M-CSFmRNAなどは確認されなかった。一方DETCでは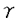 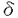 TCRmRNAが確認され、KCではIL-1,GM-CSF,M-CSFmRNAが確認されたがIL-1 mRNAはみられず、これまでの報告と一致していた。 TCRmRNAが確認され、KCではIL-1,GM-CSF,M-CSFmRNAが確認されたがIL-1 mRNAはみられず、これまでの報告と一致していた。 実験3 fLCではB7-1,B7-2分子の発現は、isotype controlと比べて229,375%の蛍光強度を示しB7-1,B7-2の発現を認めたが、CD40の発現は認めなかった。fLCを36時間培養した結果、36時間培養cLC上のB7-1,B7-2は532%,4231%と有意に蛍光強度の増加を示し、またCD40に関しても1625%の蛍光強度を示し、36時間培養cLCでCD40発現を認めた。 培養開始時にサイトカイン(M-CSF,TNF-,GM-CSF)を各々0.1ng/mlから10ng/mlまでの濃度で投与し12,36時間後のcLCのB7-1,B7-2,CD40の発現の変化を検討した。M-CSF投与では12時間後にB7-1抗原の発現増強を認めたが、B7-2,CD40発現には有意な増強は認めなかった。GM-CSF投与では、12時間でB7-1,B7-2,CD40すべての発現増強を認め、36時間でもB7-1,B7-2発現増加を認めた。TNF-投与は12時間および36時間で有意にCD40発現を増強し、36時間でB7-1の増強を誘導した。 実験4 次に免疫抑制剤(DEX,CsA,FK506)のcLCのB7-1,B7-2,CD40の発現に対する抑制作用について検討した(10-6Mから10-10Mの濃度)。DEXは、36時間培養cLCのB7-1,B7-2,CD40発現を有意に抑制した。またCsA,FK506はともに12時間培養cLCのCD40,B7-1をわずかに抑制した。B7-2への抑制作用は認めなかった。またCsA,FK506は、36時間培養cLCのCD40,B7-1,B7-2の抑制作用は示さなかった。 実験5 次にDEXにM-CSF,GM-CSFおよびTNF-を共存させた場合の変化について検討した。36時間培養LCのDEXによるB7-1の抑制作用は、TNF-投与によってほぼ完全に回復した。DEXの36時間培養cLCのB7-1,CD40の抑性作用についてはTNF-投与で変化を認めなかった。 考案 LCの抗原提示にはMHC class II抗原の発現に加えて、B7-1,B7-2を介する共刺激分子の関与、さらにCD40を介する副分子の関与が重要とされる。サイトカインによるLCのこれらの分子の発現調節については、すでにIFN-,IL-10がB7-1発現を抑制することが報告されているが、これらはbulkのECを用いた実験系であり、高純度のLCを精製した実験系は少ない。われわれは今回95%以上の高純度のLCを精製し、サイトカイン、免疫抑制剤による共刺激分子、副分子の調節について検討した。この結果、新鮮LCにM-CSFR,GM-CSFR,TNFRIIの存在を認めた。またM-CSFは12時間培養LのB7-1を増強し,GM-CSFは12時間培養cLCのB7-1,B7-2,CD40すべての発現を増強し,TNF-は12時間でB7-1を増強した。以上の結果は、LCがこれらのサイトカインに対するレセプターを有し、さらにM-CSF,GM-CSF,TNF-がLCに対して異なる抗原発現を誘導することを示している。また免疫抑制剤を用いた実験では、DEXが12,36時間培養cLCのB7-1,B7-2,CD40すべての発現を抑制し、CsA,FK-506が12時間培養cLCのB7-1,CD40の発現を軽度抑制した。これらの結果はこれらの免疫抑制剤がLCの膜抗原発現抑制に異なる作用を有することを示している。FK506のLCの共刺激分子、副分子の発現についての報告は本論文以外にはこれまで認められない。さらにDEXで誘導されたB7-1,B7-2,CD40の発現抑制に対してM-CSF,GM-CSF,TNF-の間で異なる作用のあることが確認された。以上の結果はマウスLCの共刺激分子、副分子の発現にサイトカインの関与が重要であることを示しており、今後種々の皮膚疾患におけるサイトカイン、免疫抑制剤を用いた治療の研究、開発とくにLCを用いたimmunotherapyの可能性にひとつの方向性を与えるものであると考えられる。 |
| 審査要旨 | | 本研究は、抗原提示細胞である表皮ランゲルハンス細胞(LC)表面の共刺激分子、副分子の発現に対するサイトカイン、免疫抑制剤の調節の解明を試みたものであり、マウス表皮から高純度でLCを精製し、この細胞を用いることによってM-CSF,GM-CSF,TNF-、あるいはdexamethasone、cyclosporine、FK506のLCの共刺激分子、副分子発現調節を検討し、以下の結果を得ている。またLCのレセプター発現を検討している。 1 Balb/cマウス皮膚から、表皮細胞(EC)浮遊液を作成し、EC由来のla陽性細胞を95%以上のpurityで採取した。新鮮LC(fLC)はNLDC145-,CD14-,CD11a+,CD11b+,CD11c+であり,培養LC(cLC)ではNLDC145+,CD14-,CD11a+,CD11b+,CD11c+であり、pureなLCと考えられた。 2 RT-PCR法によって、fLCにM-CSFR,GM-CSFR,TNF-RII mRNAの発現を認めた。またIL-1,TNF- mRNAなどのサイトカインmRNAの産生がfLCで確認された。しかしIL-1,GM-CSF,M-CSFmRNAなどは確認されなかった。一方DETCではTCRmRNAが確認され、KCではIL-1,GM-CSF,M-CSFmRNAが確認されたがIL-1 mRNAは認められなかった。 3 サイトカインによるLCのB7-1,B7-2,CD40発現の調節作用を以下のように解析している。%modulation of MFI(mean fluorescence intensity)=(MFI of LC treated with cytokine or immunosuppressants/MFI of LC trated with medium only)x100% fLCではB7-1,B7-2発現は、isotype controlと比べて229,375%でB7-1,,B7-2の発現を認めたが、CD40の発現は認めなかった。36時間培養のcLCのB7-1,B7-2は532%,4231%と増加を示し、またCD40に関しても1625%と増加し、36時間cLCでCD40発現を認めた。 4 M-CSF,TNF-,GM-CSFを0.1ng/mlから10ng/mlまでの濃度で投与し12,36時間のcLCのB7-1,B7-2,CD40の発現の変化を検討した。M-CSFは12時間後にB7-1の発現を増強したが、B7-2,CD40発現には変化を生じなかった。GM-CSFは、12時間cLCのB7-1,B7-2,CD40すべての発現を増強し、36時間でもB7-1,B7-2発現を増加した。TNF-は12、36時間で有意にCD40発現を増強し、36時間でB7-1の増強を誘導した。 5 免疫抑制剤(DEX,CsA,FK506)の、cLCのB7-1,B7-2,CD40の発現に対する抑制作用について検討した(10-6Mから10-10Mの濃度)。DEXは、36時間培養cLCのB7-1,B7-2,CD40発現を有意に抑制した。CsA,FK506はともに12時間培養cLCのCD40,B7-1を抑制した。B7-2への抑制作用は認めなかった。CsA,FK506は、36時間培養cLCのCD40,B7-1,B7-2の抑制作用は示さなかった。 6 DEXにM-CSF,GM-CSFおよびTNF-を共存させた場合の変化について検討した。36時間培養cLCのDEXによるB7-1の抑制作用は、TNF-投与によってほぼ完全に回復した。DEXの36時間培養cLCのB7-1,CD40の抑性作用についてはTNF-投与で変化を認めなかった。 以上の結果から、本論文はマウス皮膚由来のLCを高純度で解析し、LCの共刺激分子、副分子の発現に対してM-CSF、GM-CSF、TNF-、免疫抑制剤が異なる調節作用を有することを明かにしており、今後種々の皮膚疾患におけるサイトカイン、免疫抑制剤を用いた治療の研究、開発とくにLCを用いたimmunotherapyの可能性に方向性を与えると考えられる。以上から、本研究は学位の授与に値するものと考えられる。 |