脳腱黄色腫症(cerebrotendinous xanthomatosis:CTX)はステロール27位水酸化酵素の障害による先天性の脂質代謝異常症である。脳腱黄色腫症では、血清コレスタノール値が上昇し、腱および神経系の黄色腫、小脳症状、錐体路症状、白内障、痴呆などの症状が現れる。最初の症例はvan Bogaertらによって1937年に報告されたが、わが国では1969年の柴崎らの報告に始まる。 脳腱黄色腫症の臨床症状は、各組織にコレステロール類似物のコレスタノールが蓄積し、細胞死を引き起こすためと考え、ラットにコレスタノールを経口投与し、血清、小脳、肝臓、水晶体、眼房水などにコレスタノールが蓄積されるかどうかを検討した。次にラット小脳プルキンエ細胞、ウシ角膜内皮細胞、ウシ水晶体上皮細胞をコレスタノール添加培養液で培養し、細胞死が誘導されるかどうかを検討した。 まず5週齢のWistar系ラット(雄、体重90-110g)を1%コレスタノール含有食、1%コレステロール含有食、対照として普通食(CE-2)でそれぞれ15週間飼育した。1週毎に血清、15週後に小脳、肝臓、水晶体、眼房水を採取し、脂質(コレステロール、コレスタノール)濃度をHPLC法で測定した。6週後に小脳を摘出し、切片を作成し、Hematoxylin-Eosin染色、Sudan Black B染色、Schultz染色を行い、光学顕微鏡で観察した。ラットの体重は3群間で有意な増加を認めなかった。ラット血清コレスタノール濃度は第1週目より15週間、コレスタノール含有食群においてコレステロール食群、普通食群に比べ有意に上昇していた(p<0.05)。一方、ラット血清コレステロール濃度は6週目まではコレステロール含有食群で他の2群に比べ有意に上昇していた(p<0.05)が、その後は有意差が認められなくなった。コレスタノール含有食群では普通食群に比べコレステロール濃度が低値を示した。光学顕微鏡像では6週目の小脳スライス切片に関してHE染色で3群ともに顆粒細胞、プルキンエ細胞が同様に染色された。Sudan Black B染色ではコレスタノール含有食群ラット小脳スライス切片でプルキンエ細胞の中に薄く黒く染まる脂肪滴が見られた。次にコレステロールを特異的に染色するSchultz染色を行ったが、この脂肪滴は染色されなかった。これよりこの脂肪滴はコレスタノールの沈着と考えられた。15週後の小脳、肝臓、水晶体、眼房水コレスタノール濃度はコレスタノール含有食群(小脳:0.27±0.16mg/g tissue、肝臓:0.51±0.05mg/g tissue、水晶体:1.85±0.86mg/g tissue、眼房水:3.38±3.49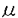 g/ml)において普通食群に比べ有意に上昇していた(p<0.05)。コレステロール濃度はコレステロール含有食群の肝臓(1.47±0.20mg/g tissue)で他の2群に比べ有意な上昇を認めた(p<0.05)。 g/ml)において普通食群に比べ有意に上昇していた(p<0.05)。コレステロール濃度はコレステロール含有食群の肝臓(1.47±0.20mg/g tissue)で他の2群に比べ有意な上昇を認めた(p<0.05)。 コレスタノール含有食ラットで小脳にコレスタノールが蓄積し、また小脳スライス切片ではプルキンエ細胞内にコレスタノールと考えられる沈着を認めたので、小脳細胞を培養しコレスタノールにより細胞死が誘導されるかどうかを検討した。胎生20-21日齢のWistar系ラット小脳をトリプシン処理により分離・播種し、D-MEM/F-12の完全無血清培地にて小脳顆粒細胞・プルキンエ細胞の初代培養を行った。6日後にエタノールに溶解した10g/mlコレステロール添加群、10g/mlコレスタノール添加群、対照として1%エタノール添加群に分けて培養を続けた。各群の培養細胞内の脂質濃度をHPLC法で測定した。TUNEL法で細胞を螢光染色し、ACAS570レーザーサイトメーターで細胞死がコレスタノールによって引き起こされるかどうかを調べた。さらにサザンブロット法でDNAラダーが生じるかどうかの解析を行った。実験培地に変更後3日目の小脳細胞(顆粒細胞とプルキンエ細胞)内の脂質濃度を測定したが、コレステロール濃度は3群間で差を認めなかった。一方、コレスタノール濃度はコレスタノール添加群(0.07±0.06g/105 cells)で他の2群に比べ有意に増加していた(P<0.05)。そこで細胞内でのコレスタノールの蓄積が小脳細胞に細胞死を引き起こしているかどうかを検討した。この細胞死がアポトーシスである可能性があると考えてTUNEL法で解析した。TUNEL法で染色される細胞を認めたので次に経時的な変化を調べた。コレスタノール添加群では3日目でTUNEL陽性細胞が37%、6日目40%と高値を示し、他の2群(ともに10%以下)と比べ有意にアポトーシスが誘導されていた(p<0.01)。培養3日目の各群の小脳細胞のDNAを抽出し、サザンブロット法で解析を行った。10g/mlコレスタノール添加群では、DNAラダーが認められアポトーシスが誘導されていることが示唆された。しかしこの培養小脳細胞は大多数が顆粒細胞でプルキンエ細胞は少数である。そこでレーザーサイトメーターで抗ラットCD-3抗体を用いプルキンエ細胞の同定を行った。これらの結果、プルキンエ細胞をコレスタノールを添加した培養液で培養すると、細胞内へのコレスタノールの取り込みが見られ、アポトーシスが誘導されると考えられる。 コレスタノール含有食ラットの水晶体・眼房水でコレスタノール濃度が上昇していたので、眼内で眼房水に接して存在する角膜内皮細胞・水晶体上皮細胞のコレスタノールによる細胞死を調べた。ウシ角膜内皮細胞・ウシ水晶体上皮細胞をトリプシン処理により分離し、10%FCS-MEM培地で各々初代培養を行った。6日後にエタノールに溶解した1,10g/mlコレステロール添加群、1,10g/mlコレスタノール添加群、対照として1%エタノール添加群に分けて培養を続けた。各群の培養細胞内の脂質濃度をHPLC法で測定した。TUNEL法にて細胞を蛍光染色し、ACAS570レーザーサイトメーターでアポトーシス誘導の経時的変化を調べた。アポトーシス断片化DNAの定量検出をApopLadder Ex/SYBR Green I法で調べた。培養12日目の角膜内皮細胞、培養24日目の水晶体上皮細胞内の脂質濃度を測定したところコレステロール濃度は3群間に有意な差を認めなかった。一方、コレスタノール濃度は角膜内皮細胞では、10g/mlコレスタノール添加群(2.01±1.53g/105 cells)で他の2群に比べ有意に上昇していた(p<0.01)。水晶体上皮細胞では10g/mlコレスタノール添加群(1.20±0.69g/105 cells)で他の2群に比べ有意に上昇していた(p<0.05)。そこで細胞内でのコレスタノールの蓄積が角膜内皮細胞、水晶体上皮細胞に細胞死を引き起こしているかどうかを検討した。この細胞死がアポトーシスである可能性があると考えてTUNEL法で経時的な変化を解析した。角膜内皮細胞では6日目はコントロール群でTUNEL陽性細胞が8%、10g/mlコレステロール添加群4%、10g/mlコレスタノール添加群6%で3群間に有意な差を認めなかったが、9日目はコントロール群8%、10g/mlコレステロール添加群14%、10g/mlコレスタノール添加群34%で10g/mlコレスタノール添加群が他の2群に比べ有意にアポトーシス誘導を認めた(p<0.05)。水晶体上皮細胞では培養10日目はコントロール群でTUNEL陽性細胞が6%、10g/mlコレステロール添加群1%、10g/mlコレスタノール添加群7%で3群間に有意な差を認めなかったが、20日目はコントロール群5%、10g/mlコレステロール添加群7%、10g/mlコレスタノール添加群27%で、また培養30日目はコントロール群7%、10g/mlコレステロール添加群20%、10g/mlコレスタノール添加群42%で10g/mlコレスタノール添加群が他の2群に比べ有意にアポトーシス誘導を認めた(p<0.05)。アポトーシス断片化DNAの定量検出を行ったところ角膜内皮細胞、水晶体上皮細胞ともに10g/mlコレスタノール添加群でアポトーシス断片化DNAが多く検出された。 以上の結果、角膜内皮細胞、水晶体上皮細胞をコレスタノールを添加した培養液で培養すると細胞内へのコレスタノールの取り込みが見られ、アポトーシスが誘導されることが示唆された。 血清コレスタノール値が上昇する脳腱黄色腫症では神経細胞や眼の細胞にコレスタノールが蓄積し、アポトーシスが誘導され、最終的に細胞死に至ると考えられる。 |