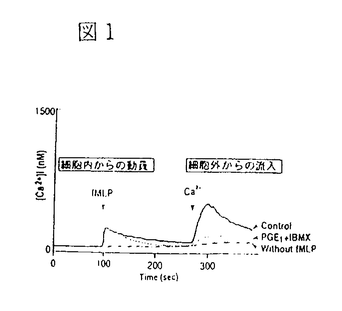
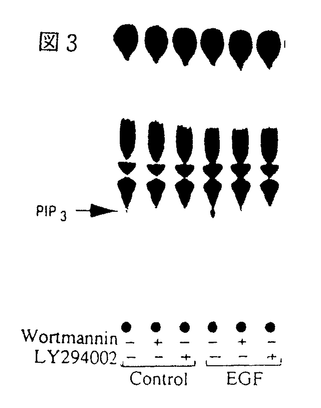
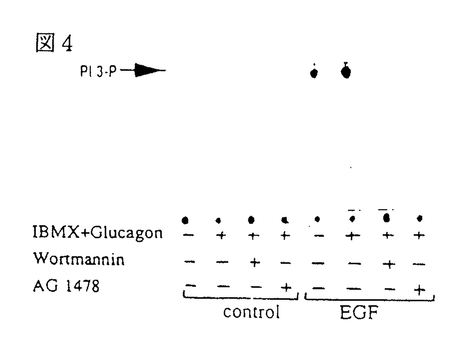
イノシトールリン脂質3-キナーゼ(PI3-K)は、イノシトール環の3位水酸基をリン酸化する脂質キナーゼで、細胞増殖因子受容体などのチロシンキナーゼ型受容体刺激により活性化される。一方、三量体G蛋白質と共役する細胞膜7回貫通型受容体を介するいくつかの細胞応答にも、PI3-Kの関与が明らかにされつつある。これらの知見は、異種のタイプの受容体刺激後のシグナル伝達経路にPI3-Kが共通して介在することを意味するが、それらの活性化と制御機構については不明の点が多い。「細胞内環状AMP産生によるホスファチジルイノシトール3-キナーゼの活性調節」と題した本論文では、先ずモルモット好中球において、G蛋白質共役型の走化性ペプチド(fMLP)受容体刺激を介する活性酸素産生が、細胞内環状AMP(cAMP)産生の増大によって抑制されることを見出し、その抑制機構について細胞内シグナル分子の産生動態から詳細に検討した結果、fMLP受容体刺激から活性酸素の産生に至るシグナル伝達経路においてPI3-Kが重要な役割を果たすこと、さらに、cAMPによる活性酸素産の抑制は、PI3-Kの活性化の抑制に作用点があることを明らかにしている。また、ラット肝の遊離細胞において、チロシンキナーゼ型受容体と結合する上皮増殖因子(EGF)の作用と細胞内cAMPの増大が与える影響を検討し、EGFによってPI3-Kが活性化されること、またこの活性化は好中球細胞の場合とは異なり、cAMP産生の増大によって逆に増強されることを見出している。 モルモット好中球おけるfMLPによる活性酸素産生とPI3-Kの活性化及びcAMP産生増大の影響 好中球にfMLPを添加すると活性酸素が放出されるが、この細胞応答は、細胞膜透過性cAMPアナログ、cAMP分解酵素の阻害剤、あるいはプロスタグランジンE1の添加によって細胞内cAMP産生を増大させると抑制された。一方、プロテインキナーゼC(PKC)を直接活性化するフォルボールエステル(PMA)も活性酸素を放出したが、この作用はcAMP産生の影響を全く受けなかった。すなわち、PKCより上流の機能蛋白質分子がcAMP依存的にリン酸化され、活性酸素産生の抑制を導くことが示唆された。PI3-Kの特異的な阻害薬であるワートマニンは、cAMP産生増大と同様に、PMAの効果に影響を与えることなくfMLPによる活性酸素産生を完全に抑制した。 fMLPによる細胞内Ca2+濃度の上昇、ホスホリパーゼA2の作用による細胞膜リン脂質からアラキドン酸放出、PI3-KによるPI-3,4,5-三リン酸(PIP3)の生成など、細胞内シグナル分子の産生に対する細胞内cAMP産生の影響と活性酸素産生との関連が詳細に検討され、fMLP受容体刺激から活性酸素の産生に至るシグナル伝達経路においては、PI3-Kが重要な役割を果たすこと、また、cAMPによる活性酸素産の抑制はPI3-Kの活性化の抑制に作用点があると考えられた。 ラット肝遊離細胞おけるEGFによるPI3-Kの活性化とcAMP産生増大の影響 ラット肝遊離細胞において、EGFはPI3-Kを活性化して細胞内PIP3産生を増大させることを見出した。EGFまたはグルカゴンで肝細胞を刺激し、細胞の抽出液を抗ホスホチロシン抗体で免疫沈降してPI3-K活性を測定した結果、EGFにより免疫沈降画分中のPI3-Kは活性化されたが、グルカゴン刺激は無効であった。しかしながら、このグルカゴン刺激によって細胞内cAMPが増大した状態においては、EGFによるPI3-Kの活性化は著しく増強された。この活性化はワートマニンとEGF受容体チロシンキナーゼ阻害薬によって抑制された。すなわち、チロシンキナーゼ型EGF受容体からPI3-Kに至る何らかの蛋白質分子がcAMP依存的にリン酸化され、EGFによるPI3-Kの活性化が増強される経路が推定された。 以上を要するに、本論文は、モルモット好中球において、G蛋白質を介したPI3-Kの活性化が細胞内cAMP産生の増大によって抑制されること、一方、肝遊離細胞における受容体型チロシンキナーゼを介したPI3-Kの活性化は、cAMPによって逆に増強されることを見出している。これらの知見は、G蛋白質共役型とチロシンキナーゼ型という異なるタイプの受容体のシグナル伝達経路に位置するPI3-Kが、細胞内cAMP産生の増大によって全く逆の制御を受けることを意味している。これらの研究成果は、今後のPI3-Kの分子多様性と生理的意義を解明する上で有益な知見であり、博士(薬学)の学位として価値あるものと認められる。 | 

 サブユニットからなる三量体G蛋白質を介してその情報を細胞内に伝達するが、このタイプの受容体刺激により惹起される種々の細胞応答にもPI3-Kの活性化が必須であることが、その強力な阻害薬であるワートマニンを用いた検討から明らかにされてきた。これらの知見は、異種のタイプの受容体刺激後のシグナル伝達経路にPI3-Kが共通して介在することを意味するが、活性化と制御機構が異なる複数のPI3-Kサブタイプの存在も指摘されている。
サブユニットからなる三量体G蛋白質を介してその情報を細胞内に伝達するが、このタイプの受容体刺激により惹起される種々の細胞応答にもPI3-Kの活性化が必須であることが、その強力な阻害薬であるワートマニンを用いた検討から明らかにされてきた。これらの知見は、異種のタイプの受容体刺激後のシグナル伝達経路にPI3-Kが共通して介在することを意味するが、活性化と制御機構が異なる複数のPI3-Kサブタイプの存在も指摘されている。