脳神経の働きは遺伝的にすべてが決定されているのではなく、生後の環境に応じて大きく変化する。この性質は「可塑性」と呼ばれ、外界の環境変化にうまく適応するのに役立つと考えられる。長期増強(LTP)は神経活動に依存したシナプス伝達可塑性の一例である。大脳皮質視覚野におけるLTPには海馬LTPには見られない「感受性期」が存在し、生後の発達段階のある特定の時期においてのみ観察される。眼が開いてしばらくの間は外界に適応するために視覚情報に対する可塑性が存在するが、ある一定の期間を経ると可塑性が消失し、獲得した性質がその後保持されるものと考えられる。このように視覚野LTPは発達に伴う可塑性の変化を研究するのによいモデルであると考えられる。 視覚野LTPの感受性期形成に関わる因子が多数想定される中で、本研究では神経伝達物質のセロトニン(5-HT)に着目した。過去には5-HTと視覚野の発達を示唆する報告例があったにも拘わらず、可塑性との関連性は明らかになっていない。本研究において、視覚野LTPに対する5-HTの作用について検討し、5-HTが感受性期を左右する因子の一つであることを見い出した。 視覚野2/3層のシナプスにおけるLTPは、白質層、視覚野4層のどちらを刺激しても誘導される。そこで、ラット(2〜6週齢)の脳から視覚野を含む切片を作成し、白質層刺激または4層刺激により誘発される集合電位を2/3層から細胞外記録し比較した。生後2〜3週齢では白質層と4層のいずれを刺激した場合もLTPが観察されたが、生後5〜6週齢では4層刺激によってのみLTPが誘導された。白質層刺激によるLTPの誘導が発達に伴い特異的に低下することが確認された。 発達に伴うLTPの変化と5-HTの関連性を探る目的で、まず、HPLCにより視覚野5-HT含有量を測定したところ、3〜6週齢で発達に伴い5-HT含有量の増加が認められた。また、5週齢ラットのスライスに5-HT受容体拮抗薬methysergideを灌流適用すると、白質層刺激によるLTPが増大した。これらの結果より、発達に伴い5-HT神経支配が増強することによってLTPが抑制される可能性が考えられたので、5-HT神経毒5,7-DHTの側脳室内投与によりLTPの低下が解除されるかどうかを検討した。5,7-DHTによる5-HT作動性神経の破壊は、抗5-HT抗体陽性細胞の消失と5-HT含有量の減少により確認された。5,7-DHT投与ラットより作成したスライスでは、5週齢であっても白質層刺激でLTPが誘導された。以上より、5-HT作動性神経の寄与が増加することによってLTPが抑制されることが示唆された。 4層刺激によるLTPは少なくとも2〜6週齢で変化しなかったので、外来性に5-HTを与えることによって4層刺激によるLTPが変化するかどうかを検討した。5-HT(10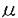 M)を灌流適用すると、通常のシナプス伝達に変化は認められなかったが、LTPの誘導がほぼ完全に抑制された。この5-HTのLTP抑制作用は濃度依存的であり(0.1-10M)、5-HT1受容体拮抗薬pindololまたは5-HT2,7受容体拮抗薬ritanserinによって拮抗されたが、5-HT3,4受容体拮抗薬MDL72222によっては拮抗されなかった。また、5-HT1A受容体作動薬8-OH-DPATと5-HT2受容体作動薬DOIもそれぞれ濃度依存的(0.1-10M)にLTPの誘導を抑制した。8-OH-DPATのLTP抑制効果はpindololで拮抗され、DOIのLTP抑制効果はritanserinにより拮抗された。以上より、5-HTが視覚野LTPを抑制することを確認し、その作用は5-HT1A、5-HT2両受容体を介することが明らかとなった。 M)を灌流適用すると、通常のシナプス伝達に変化は認められなかったが、LTPの誘導がほぼ完全に抑制された。この5-HTのLTP抑制作用は濃度依存的であり(0.1-10M)、5-HT1受容体拮抗薬pindololまたは5-HT2,7受容体拮抗薬ritanserinによって拮抗されたが、5-HT3,4受容体拮抗薬MDL72222によっては拮抗されなかった。また、5-HT1A受容体作動薬8-OH-DPATと5-HT2受容体作動薬DOIもそれぞれ濃度依存的(0.1-10M)にLTPの誘導を抑制した。8-OH-DPATのLTP抑制効果はpindololで拮抗され、DOIのLTP抑制効果はritanserinにより拮抗された。以上より、5-HTが視覚野LTPを抑制することを確認し、その作用は5-HT1A、5-HT2両受容体を介することが明らかとなった。 次に、5-HTのLTP抑制作用にGABAを神経伝達物質とする抑制神経系が関与する可能性について検討した。スライスにGABAA受容体拮抗薬bicucullineを灌流適用しGABA作動性神経を遮断した条件下でも、8-OH-DPATはLTPを抑制したが、DOIはLTPを抑制しなかった。以上の結果より、5-HT1A、5-HT2受容体を介したLTP抑制にはそれぞれ異なった機序が関与していること、特に5-HT2受容体を介したLTP抑制はGABA作動性神経を介することが示唆された。 以上の結果より、5-HTは視覚野LTPの感受性期の終結を決定する因子の一つであると考えられた。本研究は、特定の神経伝達物質が発達に伴う可塑性の消失に関与することを示した例であり、今後の脳の可塑性の研究への貢献が大きく、博士(薬学)に値すると判断した。 | 



