近年、心臓薬物治療における延命効果に心拍数調節の重要性が明らかになってきた。心拍数調節には心臓洞房結節の複数のチャネルが寄与しており、その中の一つに過分極活性型のIfチャネルがある。Ifチャネルは自発的に発生する洞房結節の活動電位の緩徐脱分極相の傾きを形成しており、この傾きは心拍数を決める要因の1つである。自律神経ホルモンによる心拍数調節には、受容体刺激によりIfチャネルが細胞内cAMPにより直接調節されることが主に寄与していると考えられている。しかし、Ifチャネルはいまだクローニングがなされておらず分子構造又はcAMP結合部位に関する情報は全くない。特に生理学的な状態でのムスカリン性の心拍数制御については、細胞内cAMP低下によるIfチャネル電流(If)抑制が主に関与していると考えられているが、その細胞内制御機構を明らかにした研究はほとんどない。また、Ifチャネルは細胞内の重要なメッセンジャーであるCa2+によって活性化されるが、その機構は不明である。 本論文の目的は、洞房結節細胞において、細胞内cAMPのIfチャネル制御機構を調べることにより自律神経による心拍数調節のメカニズムを理解することと、細胞内Ca2+よるIfチャネル制御機構を調べることにより新たな制御経路を見い出すことである。まず、Ifチャネルの細胞内cAMP調節部位の位置がカルボキシル末端に存在するか調べるために、洞房結節細胞内に蛋白分解酵素を投与し、細胞内cAMPによるIfチャネル調節への効果を検討した。また、Ifチャネルのムスカリン性刺激による細胞内cAMP制御機構を明らかにするために、ムスカリン性If抑制作用に対するホスホジエステラーゼ(PDE)阻害剤とNO合成酵素阻害剤の作用を調べた。さらに、細胞内Ca2+によるIfチャネル制御に対するcalmodulinの関与をcalmodulin antagonistの投与により検討した。 ウサギ心臓を酵素的に消化し、単離した洞房結節細胞を用い、whole-cell patch-clamp法で膜電流を測定した。Ifチャネルの制御へのカルボキシル末端の関与を調べるために、カルボキシル末端からアミノ酸を順々に切断する酵素(carboxypeptidase A)を洞房結節細胞内に投与し、Ifに対するl-isoproterenol(Iso)とcarbachol(CCh)の作用を調べた。その結果、carboxypeptidase Aの細胞内投与により、IsoとCChの作用が消失することを見出し、酵素がcAMP調節部位を切断している可能性が考えられた。次に、細胞内に投与した酵素が作用していることの指標として、Ifと同時にL型Ca2+電流(ICa(L))を同一細胞で測定しcAMPの作用を調べた。酵素の細胞内灌流により、ICa(L)は以前の報告通りの作用を示し、それと同時にIfは減少し、cAMPへの感受性の消失が見られた。さらに、酵素の細胞内灌流により、細胞膜破壊後の時間変化に伴うIfチャネル活性化の電位依存性が過分極側へシフトし、さらに活性化の電位依存性のcAMP感受性は消失した。その結果、Ifチャネルの細胞内カルボキシル末端がチャネル活性に関与し、cAMP調節部位が細胞内カルボキシル末端に位置することを見い出した。 次に、非選択的PDE阻害剤であるIBMXとcGMP依存性PDE阻害剤であるEHNA、それぞれの存在下でムスカリン受容体アゴニストであるCChによるIf抑制作用が減弱したことから、Ifチャネルのムスカリン性制御にcGMP依存性PDE typeIIが関与していることを明らかにした。また、NO合成酵素阻害剤(L-NMMA)を処置したところ、CChによるICa(L)抑制作用は有意に減弱したが、If抑制作用は影響を受けなかった。よって、L型Ca2+チャネルとは異なり、Ifチャネルのムスカリン性制御にはNO合成系の関与がないことが明らかになった。今回の結果により、洞房結節細胞におけるIfチャネルのムスカリン性制御は、NO合成系以外の経路でcGMP依存性PDE typeIIの活性化を介することを初めて示した。 さらに、Ifチャネルの細胞内Ca2+による機能制御における機能蛋白の関与を調べるために、Ifに対するcalmodulin antagonistの作用を検討した。W7とcalmidazoliumという2種のcalmodulin antagonistがIfを抑制したことから、Ifチャネルの細胞内Ca2+による機能制御にcalmodulinが関与すると考えられる。 以上、本研究は、ウサギ洞房結節細胞のIfチャネルにおいて、1)cAMP調節部位が細胞内カルボキシル末端に位置すること、2)ムスカリン刺激による細胞内cAMP調節にNO合成系の関与がなくcGMP依存性ホスホジエステラーゼが関与すること、3)細胞内Ca2+による機能制御にcalmodulinが関与していることを示した。本研究で得られた知見は、細胞内cAMPによる調節という機能をIfチャネルの分子構造と初めて結びつけ、Ifチャネルのムスカリン性制御の新たな経路を示すことで自律神経ホルモンによる心拍調節機構の解明の糸口を与えるものであり、Ca2+によるIfチャネルの細胞内制御に新しい蛋白の関与を示唆するものである。本研究は、心拍調節の生理学および薬理学に貢献をするものであり、博士(薬学)の学位に値するものと認めた。 |  アドレナリン受容体刺激で細胞内cAMP量が増加すると、活性化の電位依存性が脱分極側にシフトして電流が増大することが明らかにされている。
アドレナリン受容体刺激で細胞内cAMP量が増加すると、活性化の電位依存性が脱分極側にシフトして電流が増大することが明らかにされている。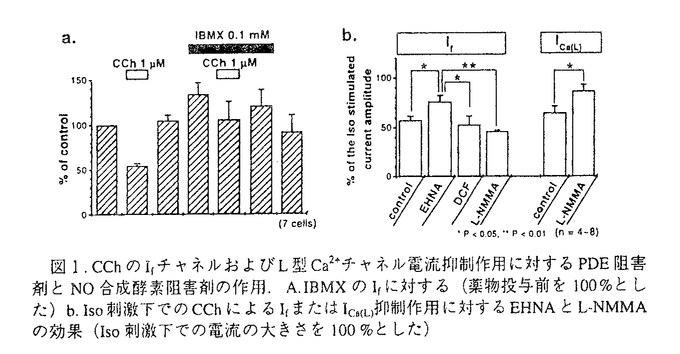

 M)とCCh(1
M)とCCh(1