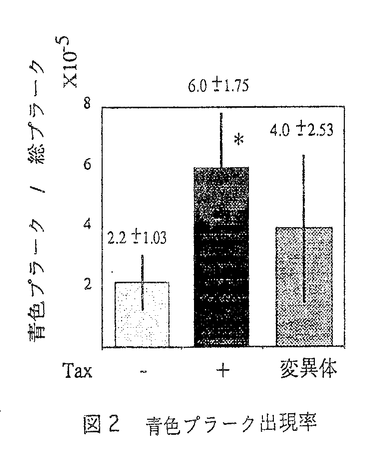
成人T細胞白血病(ATL)は、ヒトレトロウイルスHTLV-1の感染が原因で発症する特異な白血病として知られている。このウイルスによる白血病発症のメカニズムとして、ウイルスの制御遺伝子であるTaxを中心に、分子生物学的研究が進められ、その初期段階について多くの理解が得られている。しかしながら、がんは多段階の遺伝子異常の蓄積により発症する遺伝子病であることは、広く受け入れられているところである。従って、感染したウイルスの遺伝子が、直接細胞の機能を制御することに加えて、その後に引き続いておきると予想されるいくつかの遺伝子変化の解明が重要になるが、それらの実体と異常の発生機構については、未だ推論の域を出ていない。 本論文は、この点に焦点を当て、HTLV-1の遺伝子発現が、細胞の遺伝子の突然変異を促進することを証明したものである。ATL細胞が高頻度の染色体異常を持ち、ゲノムの不安定性をきたしていると考えられることから、細胞の突然変異との関連に興味を持ち、ATLの発症に重要な鍵を握ると予想されているTaxに注目した。まず、Taxを発現した細胞では、6-メルカプトプリン(6MP)にたいする耐性細胞の出現が高いことから、Hypoxantine guanine phosphoribosyl transferase(HPRT)の突然変異を促進する可能性を示した。この現象を塩基配列のレベルで確実に証明するために、細胞の染色体DNAに、大腸菌のLacZ遺伝子の発現を制御するLacI遺伝子を組み込んだ細胞を用い、PCRを使用することなく、多数の細胞におけるLacIゲノム遺伝子の変異をスクリーニングするアッセイ系を導入した。このアッセイ系では、一定の条件にさらした後にパッケイジングによって再生されるレポーターファージに変異が入っておれば、特定のプレート上で青色を呈するので、多くのファージをスクリーニングすることが可能である。この方式を用いて、HTLV-1のTaxを発現した細胞では、LacIの突然変異率がTaxを発現しない場合に比べて3倍に上昇することを示した。さらに、LadI遺伝子上の変異の位置、その種類を塩基配列の決定により解析した結果、変異のおきる位置およびその種類には特徴がなく、ランダムであることも明らかにした。 このことは、Taxが、細胞の転写因子や細胞周期を調節する特異的な遺伝子の修飾を介して、細胞の異常増殖を促進するほかに、細胞遺伝子の変異を促進して間接的に細胞の増殖や機能に異常をもたらす頻度を上昇させることを証明したもので、高く評価される。即ち、HTLV-1は、感染細胞の初期段階における異常増殖のみでなく、それに引き続いておきると考えられている細胞遺伝子の変異(異常)の誘導にも関わることを証明したものである。その変異率亢進の機構についての解析は多くは進まなかったが、Taxが特定の塩基あるいは反復配列に特異性を持つDNA修復遺伝子などではなく、より一般的な反応に関わる分子を介して突然変異率を上昇させることが推論された。例えば、DNAポリメラーゼ の発現がTaxにより抑制されることは既に報告されているが、その結果としてランダムな変異率が上昇する説明が可能である。しかし、実際にHTLV-1感染細胞の変異率が真に高いかどうかについての実証もなかった。このようなランダムな変異率の上昇により、感染細胞がより高頻度に変異を繰り返し、ついにはがん細胞として異常増殖する可能性を強く支持する。 の発現がTaxにより抑制されることは既に報告されているが、その結果としてランダムな変異率が上昇する説明が可能である。しかし、実際にHTLV-1感染細胞の変異率が真に高いかどうかについての実証もなかった。このようなランダムな変異率の上昇により、感染細胞がより高頻度に変異を繰り返し、ついにはがん細胞として異常増殖する可能性を強く支持する。 以上、本研究は、HTLV-1による発がん機構に、今まで知られていない新しい視点を事実をもって示したものである。特に単一のウイルス蛋白が、極めてカテゴリーを異にする細胞機能に、無差別に影響を与えうることを証明した観点からは、HTLV-1のみならず他のウイルスによる多段階発がんの研究に対しても、大きく寄与するものであり、博士(薬学)の学位に相当すると判断される。 |