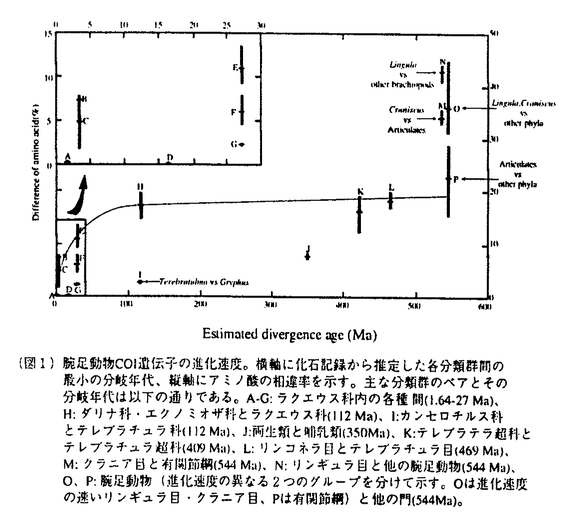
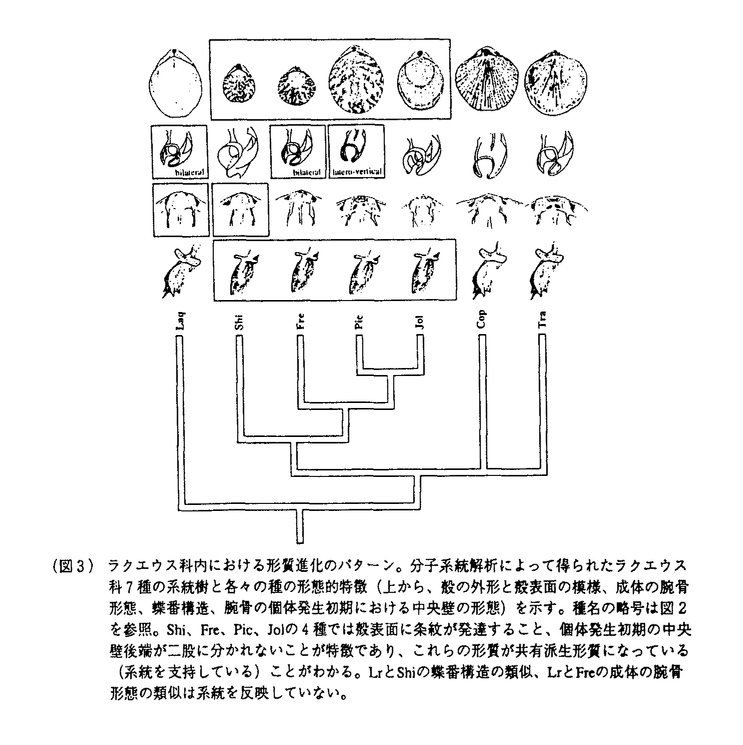
| 審査要旨 | | 本論文は第一部,第二部の2つの部分からなる.第一部では,腕足動物全般におけるチトクロムcオキシダーゼサブユニットI(COI)遺伝子の進化速度と進化様式,およびその分子系統学的有用性について主に述べられており,第二部では,これらの知見をふまえた上で行われた,COI遺伝子の塩基配列の比較による腕足動物ラクエウス科の系統解析の結果とその意義についてまとめられている. COI遺伝子は,さまざまな動物分類群の分子系統解析のマーカーとして多用されているが,その際COI遺伝子の進化速度や進化様式について知っておくことが前提となる.そのための信頼できる唯一の方法は,分子の分岐年代を化石記録の出現,大陸の分裂,海洋の分断などの地質学的な証拠から推定し,これらの年代をもとに分子の一次配列の比較をすることである.しかし,これまでは一部の脊椎動物における研究があるのみで,化石記録が豊富な海棲無脊椎動物を用いた研究はなされていなかった.その意味で,本論文の第一部で述べられている研究結果は重要であり,これまで少ないデータから推測するしかなかったCOI遺伝子の進化速度と進化様式を,より明確にしたという点で高く評価できる. 腕足動物は,顕生代を通じて連続的な化石記録を有する数少ない動物門の一つで,特に古生層の生層序学的研究に古くから多用され,化石分類群の出現年代について綿密なデータが蓄積されている.また,古生代に成立したものを含む腕足動物の主要な分類群は,現在も世界中の海底に生息しているため,それぞれからDNAを抽出して配列の比較を行うことができる.本論文の第一部で述べられている研究では,腕足動物の現存する5つの目(リンギュラ目,クラニア目,リンコネラ目,テシデア目,テレブラチュラ目)すべてをカバーする20種32個体のCOI遺伝子の大半の領域(1218塩基対)の塩基配列を決定し,それぞれの種間の配列の変異と化石記録から推定された分岐年代との比較を行った. その結果,大部分の腕足動物では,約1億年前以前に分岐した分子間では,塩基置換,アミノ酸置換ともに飽和に達していること,また系統によっては,進化速度の著しく速いものと遅いものもあること,そしてこれらの進化速度の違いは塩基組成の違いと相関があることが判明した.さらに得られた配列の系統解析の結果から,約1億年前より以前に分岐した分類群間の系統推定には,同一座位の多重置換によるノイズの影響を考慮する必要があることを示した.また本研究では,COIの配列進化の保守性を利用して遺伝暗号の推定も行っており,腕足動物におけるミトコンドリアDNAの変則暗号を初めて明らかにしている. 本論文の第二部では,第一部の結果から信頼のおける分子系統推定のできることが確認された(すなわち約1億年前以降に放散した)テレブラチュラ目ラクエウス科の7属12種を用いて,得られた分子系統樹をもとに形態形質の進化過程を推察した結果を論じている.調べた形質は,触手冠(摂食・呼吸器官)を支える腕骨と呼ばれる石灰質の骨格の形態,殻の外形,表面装飾,蝶番構造などであり,これらのうち,個体発生初期における腕骨の形態と殻表面の模様が系統を反映していることを明らかにした.一方,成体の腕骨の形態や殻の外形,蝶番構造の特徴などは必ずしも系統を反映しておらず,従来行われてきた化石も含めた系統推定の多くが再検討を要することを論じている. デボン紀に成立し,新生代に大きく放散した腕足動物のテレブラチュラ目は,腕骨を持つことで特徴づけられる.テレブラチュラ目の進化史は,腕骨形態の進化史とほとんど同義であったとも言えるが,実は,腕骨形態はテレブラチュラ目の最も重要な分類形質と考えられてきており,従来の腕骨形態に基づく進化史の議論は,宿命的に循環論であった.本論文第二部で述べられているアプローチは,形態とは独立の分子の形質から系統を推定し,形態の進化過程を推定するというものであり,化石も含めた形態進化の研究に新しい方向性を示したという意味で意義深い. なお,本論文第一部は小島茂明博士,遠藤一佳博士との,本論文第二部は遠藤一佳博士との共同研究であるが,論文提出者が主体となって実験,解析および検証を行ったもので,論文提出者の寄与が十分であると判断する.本論文は,化石記録の利点を十分に生かし,分子系統学に大きく寄与した点,そして分子データと初期発生の観察をもとに化石記録を見直し,形態進化過程を考察する,という進化古生物学の新しい領域を開拓した点で傑出したものである.よって審査委員全員は,論文提出者に博士(理学)の学位を授与できると判断した. |