本研究はPAF受容体のリガンド結合とシグナル伝達を解明する目的でCHO細胞にPAF受容体を安定発現し、PAF受容体の構造、PAF受容体の細胞内移行について解析し、下記の結果を得ている。 1.モルモット、ヒト、ラット、マウスの4動物種のPAF受容体の[3H]PAF結合のスキャッチャード解析ではいずれにおいても直線に回帰され、リガンドの結合部位は一つと考えられた。アラキドン酸の放出は4種ともPAF0.1nMでみられ、10nMで最大に達した。また、4種のPAF受容体において亜鉛イオンによりリガンド結合が阻害された。リガンド結合は亜鉛イオンによりBmaxをほとんど変えずにKd値を上昇させたことより、リガンド結合に対する亜鉛イオンの阻害作用は競合阻害と考えられた。この結果より亜鉛イオンの配位し得ると考えられる3つのヒスチジンの部位がPAF結合に重要な部位である可能性が示された。PAFによるアラキドン酸の放出についても亜鉛イオン濃度依存性に阻害され、亜鉛イオンによるリガンド結合の阻害が、シグナル伝達に関わることが示された。亜鉛イオンによりリガンド結合が阻害されることより、PAF受容体へのPAFの結合部位に3つのヒスチジン残基(His-188,His-248,His-249)が関与している可能性が示され、以前提唱されたモデルを支持する結果が得られた。 2.PAF受容体の細胞内移行の有無とその性質を明らかにする目的でモルモットPAF受容体の細胞外アミノ末端に標識ペプチド(tag)を導入する変異を行い、細胞表面への抗tag抗体結合量を測定することにより細胞膜受容体量を測定してその動態を解析し、蛍光顕微鏡による観察も併せて行っている。アゴニスト刺激によりPAF受容体は、アゴニスト濃度依存的、時間依存的に細胞内へ移行した。この移行はクラスリン被覆小胞の形成を阻害する各種前処理によりほぼ完全に阻害され、クラスリンを介した経路によると考えられた。 3.PAF受容体は細胞内移行後、PAF受容体のアンタゴニストである、WEB2086、TCV-309によるアゴニスト除去により細胞膜に再分布することが観察された。蛋白合成阻害剤のシクロヘキシミドによりこの再分布は阻害されず、再分布した受容体は細胞内移行した受容体のリサイクルと考えられた。再分布した受容体にPAF刺激をしたところ、再度細胞内移行が観察された。 以上本論文は、PAF受容体へのPAFの結合部位に3つのヒスチジン残基(His-188,His-248,His-249)が関与している可能性が示し、リガンド刺激によるクラスリン依存性のPAF受容体の細胞内移行を初めて明らかにした。さらにPAF受容体はアンタゴニストによるアゴニスト除去によりリサイクルし、再感作されていた。PAF受容体のリガンド結合とシグナル伝達に関して、PAF受容体の亜鉛イオンによる阻害の解析、およびリガンド依存性細胞内移行のメカニズムとその意義に注目し、新しい知見を得ており、学位の授与に値するものと考えられる。 | 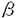 2アドレナリン受容体で解析が進んでおり、アゴニスト刺激により受容体の細胞内カルボキシ末端がリン酸化され、このリン酸化部位に結合する蛋白質と細胞膜被覆小胞の構成成分であるクラスリンとの相互作用により、細胞内移行することが報告されている。受容体の細胞内移行は脱感作の機序の一部と考えられていたが、脱感作には受容体のリン酸化の方が迅速であり、むしろ細胞内移行は受容体のリサイクルや再感作のために重要な役割を果たしていると考えられている。
2アドレナリン受容体で解析が進んでおり、アゴニスト刺激により受容体の細胞内カルボキシ末端がリン酸化され、このリン酸化部位に結合する蛋白質と細胞膜被覆小胞の構成成分であるクラスリンとの相互作用により、細胞内移行することが報告されている。受容体の細胞内移行は脱感作の機序の一部と考えられていたが、脱感作には受容体のリン酸化の方が迅速であり、むしろ細胞内移行は受容体のリサイクルや再感作のために重要な役割を果たしていると考えられている。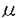 Mに達すると言われている。PAF受容体が海馬の神経細胞に存在することを考えると、亜鉛はPAFの神経作用を調節している可能性も考えられる。
Mに達すると言われている。PAF受容体が海馬の神経細胞に存在することを考えると、亜鉛はPAFの神経作用を調節している可能性も考えられる。