種々の細胞膜7回貫通型受容体は、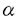   サブユニットからなる三量体Gタンパク質を介して細胞内にシグナル分子を生産する酵素やイオンチャネルにその情報を伝達している。アゴニストがこのタイプの受容体に結合すると、Gタンパク質はGTPの結合したサブユニットとサブユニットに解離するが、近年サブユニットに加えて、サブユニット(G)もいくつかのエフェクター分子の活性を直接制御することが見出されてきた。Gには、一次構造上約40アミノ酸単位からなるWDモチーフと呼ばれるユニークな配列(シート→ターン→シート→ターンと続く)が7回繰り返して存在している。最近のX線構造解析から、GはN末端側からのヘリックスに続いて、WDモチーフの1つがプロペラの羽根構造を形成し、全体として7枚羽根のプロペラー様構造をとるように配置されていることが示された(図1)。 サブユニットからなる三量体Gタンパク質を介して細胞内にシグナル分子を生産する酵素やイオンチャネルにその情報を伝達している。アゴニストがこのタイプの受容体に結合すると、Gタンパク質はGTPの結合したサブユニットとサブユニットに解離するが、近年サブユニットに加えて、サブユニット(G)もいくつかのエフェクター分子の活性を直接制御することが見出されてきた。Gには、一次構造上約40アミノ酸単位からなるWDモチーフと呼ばれるユニークな配列(シート→ターン→シート→ターンと続く)が7回繰り返して存在している。最近のX線構造解析から、GはN末端側からのヘリックスに続いて、WDモチーフの1つがプロペラの羽根構造を形成し、全体として7枚羽根のプロペラー様構造をとるように配置されていることが示された(図1)。  図1 G-サブユニットのプロペラ様構造 図1 G-サブユニットのプロペラ様構造 そこで私は、Gの構造と機能をより詳細に解析する目的から、動物細胞でのインタクトなG複合体の発現系を確立し、GのWDモチーフに変異を導入した種々の組換え体を作製してGで直接活性化される酵素に与える影響を異なる手法で検討した。その結果、G-エフェクター間の結合において、Gの7つのWDモチーフがエフェクターの種類によって使い分けられていることを見出した。さらに、Gで活性化されるイノシトールリン脂質3キナーゼ(PI3K)側の機能部位についても検討を加えた。 1.COS-7培養細胞におけるG複合体の発現:発現Gの性状解析 2種の異なるサブユニットからなり、そのサブユニットがイソプレニル化されるGの発現系としては、バキュロウイルスなどが利用されてきたが、多種の変異体の調製には必ずしも適当な系とは考えられない。そこで私は、エレクトロポレーション法を用いてCOS-7細胞にタグ付きGとGのcDNAを同時に導入し、Gを発現したCOS-7細胞からタグを利用してGを精製した。さらに脂質ミセルによって沈降濃縮させる手法を用いて、比較的短時間で高濃度のG標品を得ることに成功した。 得られたタグ付きGがインタクトな構造でありサブユニットと会合し得ることは、百日咳毒素によるGサブユニットのADPリボシル化で確認した。Gで活性化されることが知られているヘテロダイマー型(p110/P85)のイノシトールリン脂質3キナーゼ(PI3K)にこのG標品を添加してPIP3産生量を測定した結果、確かにPI3Kの活性化が認められた。 2.Gによるアデニル酸シクラーゼの活性化:各種のGキメラ体を用いた解析 GのWDモチーフの役割を検討するために、1〜7のそれぞれのWDモチーフのコア領域をホスフォリパーゼA活性化タンパク質の第5WDモチーフに置換した遺伝子を構築した(図2)。2型のアデニル酸シクラーゼは、GSのサブユニットによって活性化され、サブユニットによってその活性がさらに増強される特性をもつことが知られている。そこでアデニル酸シクラーゼに対するGのWDモチーフ相互作用部位を検討するために、COS-7細胞にアデニル酸シクラーゼ(2型)のcDNAと、タグ付きのGまたはGキメラ体のcDNA、さらにGのcDNAを同時に導入した。導入COS-7細胞を受容体アゴニストのイソプロテレノールで刺激してGSのサブユニットを解離させ、各種のGキメラ体による増強効果を検討した。その結果、図3に示すように、アデニル酸シクラーゼ(2型)はGの第1WDモチーフを中心として相互作用していることが示された。  図2 G2とPLA活性化タンパク質とのキメラ体 図2 G2とPLA活性化タンパク質とのキメラ体 図3 Gキメラ体による2型アデニル酸シクラーゼの活性化:第1WDモチーフの関与3.GとRaf-1キナーゼとの会合:酵母のtwo-hybrid系を用いた解析 図3 Gキメラ体による2型アデニル酸シクラーゼの活性化:第1WDモチーフの関与3.GとRaf-1キナーゼとの会合:酵母のtwo-hybrid系を用いた解析 Raf-1キナーゼはCキナーゼやRasを介したMAPキナーゼカスケードにおい重要な役割を担うセリン/スレオニンキナーゼであるが、その活性化経路には不明な点も残されている。膜移行シグナルを付加した実験等からRaf-1の細胞膜への移行が活性化に重要であると考えられているが、この点と関連して、最近Raf-1キナーゼのN末端調節領域とGのC末端部位との結合が、酵母のtwo-hybrid系あるいは精製タンパク質を用いた系で報告された。そこで私は、このRaf-1キナーゼに対するG上の結合部位を調べるために、先ずC末端側3単位からなるWDモチーフを2種類作製し、Raf-1キナーゼのN末端領域との相互作用を酵母のtwo-hybrid系を用いて検討した。その結果、図4に示すように、第4〜7WDモチーフ間で相互作用することが確認できた。さらにその部位を特定する目的で、1単位のみからなる各種のWDモチーフを作製して検討を加えた結果、Raf-1キナーゼはC末端側の第6WDモチーフ以外に、第3WDモチーフとも相互作用する可能性が新たに見出された。  図4 酵母のTwo-hybrid系を用いたGとRaf-1との結合部位の解析4.Gによるイノシトールリン脂質3キナーゼ(PI3K)の活性化 図4 酵母のTwo-hybrid系を用いたGとRaf-1との結合部位の解析4.Gによるイノシトールリン脂質3キナーゼ(PI3K)の活性化 PI3KはPI,PI(4)P,PI(4,5)P2を基質として、そのイノシトール環3位をリン酸化する酵素であり、細胞の分化、増殖などに重要な役割を果たす脂質キナーゼである。PI3Kにはアイソザイムが存在するが、当教室ではヘテロダイマー型のPI3K(p110/P85)がGとチロシンリン酸化ペプチドによって相乗的に活性化されることを見出している。そこで、調節サブユニットp85の一部を付加した恒常的活性化型p110とp110の変異体をそれぞれTHP-1細胞に遺伝子導入し(図5)、受容体刺激によるPI3Kの活性化を無傷細胞のレベルで検討した。その結果、活性化型p110を発現した時に、Gタンパク質共役型fMLP受容体の刺激で細胞内にPIP3産生が観察され、Gは調節サブユニットであるp85側ではなく、触媒サブユニットであるp110に作用することが示された。  図5 p110-PI3K遺伝子導入によるfMLP受容体刺激を介する細胞内PIP産生の増大5.まとめ 図5 p110-PI3K遺伝子導入によるfMLP受容体刺激を介する細胞内PIP産生の増大5.まとめ 本研究において私は、GのWDモチーフに変異を導入した種々の組換え体を用いて、アデニル酸シクラーゼ、Raf-1キナーゼに対する影響を検討し、G-エフェクター間の結合においては、Gの7つのWDモチーフがエフェクターの種類によって使い分けられていることを見出した。さらに、Gで活性化されるイノシトールリン脂質3キナーゼ(PI3K)についても検討を加え、GがPI3Kの触媒サブユニットであるp110上の機能部位に作用することを見出した。 | 
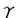 サブユニットとその標的タンパク質の機能部位に関わる解析
サブユニットとその標的タンパク質の機能部位に関わる解析