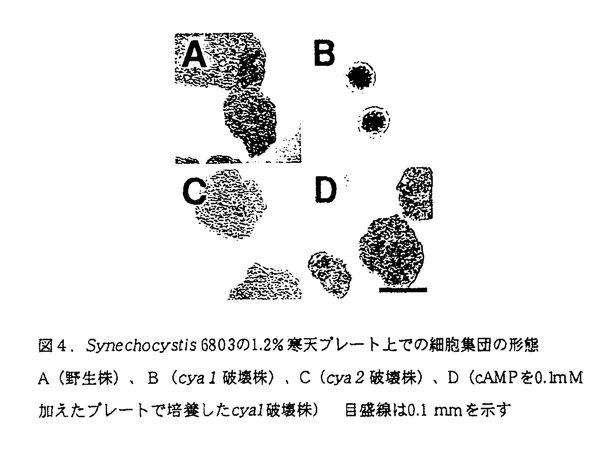
サイクリックAMP(cAMP)は原核生物から真核生物まで広く存在する細胞内情報伝達物質であり、アデニル酸シクラーゼによってATPから合成される。アデニル酸シクラーゼ遺伝子はラン藻から多数単離されているが、これまで外部環境変化に対応する個々のアデニル酸シクラーゼの役割やcAMPの具体的な機能については全く解明されていない。 申請者は、ラン藻Synechocystis sp.PCC6803において、青色光-cAMP情報伝達機構の存在を明らかにし、その中でのアデニル酸シクラーゼの役割について解析した。 初めに、Synechocystis6803を用い、光照射後数分以内に細胞内cAMP量が、暗所での細胞内cAMP量の数倍にまで増加することが明らかにした。さらに、この反応に関与する光質を調べるために、岡崎国立共同研究機構基礎生物学研究所の大型スペクトログラフを用いて、単色光照射による細胞内cAMP量の変化を調べた。その結果、青色光(450 nm)の照射によりcAMP量が増加し、520nm以上の光ではcAMP量は変化しないことが明らかとなった。また、光によるcAMPの増加は、フラビンの光反応の阻害剤であるフェニル酢酸により阻害されたことから、フラビンが関わっていることが示唆された。 Synechocystis6803のゲノムの全塩基配列は1996年に決定されている。その中に、アデニル酸シクラーゼと考えられるオープンリーディングフレーム(ORF)が2つ存在した。これらのORFをcya1、cya2と名付けた。どちらのアデニル酸シクラーゼが青色光に対する反応に関わっているかを明らかにするために、Synechocystis6803のcya1、cya2破壊株を作製し、暗から明への光環境の変化にともなうcAMP量の変化を調べた。cya2破壊株では、野生株と同様に、青色光照射によりcAMP量が増加したが、cya1破壊株では青色光の照射によるcAMP量の増加はみられなかった。このことから、野生株でみられた青色光によるcAMP量の増加は、アデニル酸シクラーゼCya1の活性化が青色光により引き起こされたためであると考えられる。 さらに、cya1破壊株には興味深い表現型がみられた。通常Synechocystis6803の野生株は、運動性をもつことが知られており、プレート上でシート状にひろがってコロニーを形成しない。しかしcya1破壊株はプレート上でコロニーを形成し、運動性が失われていると考えられる。cya2破壊株は、野生株と同様の表現型を示した。これらの株の細胞内cAMP量をエンザイムイムノアッセイ法により測定した。cya1破壊株でのみ細胞内cAMP量の顕著な減少がみられた。また、培地中にcAMP(0.1mM)を加えた寒天プレート上では、cya1破壊株は野生株と同じようにコロニーを形成しなくなり、cAMPの添加によりcya1破壊株細胞の運動性が回復したと考えられる。以上の結果からSynechocystis6803の運動にはcAMPが必要であることが示唆された。さらに、Synechocystis6803の細胞の運動を顕微鏡下で観察し、ビデオ録画して解析した。細胞に青色光(450nm)、赤色光(670nm)を照射して運動能を検討した結果、青色光がもっとも顕著に細胞の運動を促進した。cya1破壊株は青色光を照射しても運動性を示さなかった。 以上の結果より、Synechocystis6803では、青色光のシグナルがCya1に伝わり、Cya1の活性化により細胞内cAMP量が増加し、それによって細胞の運動が促進されること、すなわち青色光-cAMP情報伝達機構が存在し、それが細胞の運動を調節していることを明らかにした。本研究はラン藻のアデニル酸シクラーゼの具体的な生理機能を初めて明らかにしたものであり、同時にラン藻の運動の制御に関する分子生物学的な研究の出発点となる新しい基礎的な知見を提供し得た。 上記のように、申請者は分子生物学的手法を用いて生理学的研究を発展させる幅広い知識と研究能力を身につけてきたと考える。また、ラン藻を材料としたcAMP情報伝達に関する研究において、独創性を示し、この分野の発展に多大の貢献をした。よって、本論文は博士(学術)の学位請求論文として合格と認められる。 |  mol/m2/s)を照射し、光照射の前後に細胞懸濁液を採取した。cAMPはエンザイムイムノアッセイ法により測定した。その結果、光照射後数分以内に細胞内cAMP量が、暗所での細胞内cAMP量(約40 pmol/mg chl.)の数倍にまで増加することが明らかになった。
mol/m2/s)を照射し、光照射の前後に細胞懸濁液を採取した。cAMPはエンザイムイムノアッセイ法により測定した。その結果、光照射後数分以内に細胞内cAMP量が、暗所での細胞内cAMP量(約40 pmol/mg chl.)の数倍にまで増加することが明らかになった。