zif268はDNA結合部位に3つのzinc fingerを持つ転写因子をコードする前初期遺伝子である。zif268遺伝子は様々な細胞や組織において、様々な刺激により速やかに発現することが示されており、細胞の特性を長期に渡り変化させる「第三のメッセンジャー」として機能すると考えられる。特に、zif268のmRNAのラット海馬における発現は、記憶・学習のモデルと思われるlong term potentiation(LTP)の形成と相関性があることがわかっており、このzif268遺伝子の発現が神経細胞内でどの様なシグナル伝達を介して発現するのかを解析することは、記憶・学習の形成のメカニズムの分子レベルでの解明に役立つと考えられる。そこで、神経細胞のモデル細胞であるPC12D細胞を用いて、様々な刺激によるzif268遺伝子の発現を制御するシグナル伝達系の解析を行った。 PC12D細胞は膜上に高親和性NGFレセプターであるtrkA及びムスカリン性アセチルコリンレセプター(mAChR)のM1及びM4サブタイプを発現している。PC12D細胞をNGF或いはmAChRアゴニストであるカルバコールで刺激するとzif268mRNAレベルが一過性に上昇する。NGF刺激の場合、このmRNAレベルの上昇に先立ちmitogen-activated protein kinase(MAPK)が活性化されることがわかっている。また、カルバコール刺激の場合、MAPK及びc-Jun N-terminal kinase(JNK)が強く活性化されることが示されている。 MAPK及びJNKはスレオニン及びチロシン残基のリン酸化により活性化されるセリン・スレオニンキナーゼであり、p38MAPKと共にMAPKスーパーファミリーを形成している。MAPKは様々な刺激により活性化されるが、特にレセプター型チロシンキナーゼの活性化により活性化される経路(MAPKカスケード)について、レセプターのチロシン残基のリン酸化を介してShc/Grb2,mSOS,Ras,Raf-1或いはB-Raf,MAPKと至る経路が同定されている。また、Gタンパク質共役受容体の活性化によるMAPKの活性化についても、protein kinase C(PKC)及びRaf-1を介する経路、細胞内カルシウムの上昇からチロシンキナーゼPYK2及びRasを介する経路、また、Gタンパク質の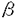  サブユニット及びRasを介する経路などが同定されている。MAPKは活性化されると核内に移行し、serum response factor(SRF)と結合して遺伝子発現を促進するternary complex factor(TCF)をリン酸化し活性化することにより、遺伝子発現を促進する。 サブユニット及びRasを介する経路などが同定されている。MAPKは活性化されると核内に移行し、serum response factor(SRF)と結合して遺伝子発現を促進するternary complex factor(TCF)をリン酸化し活性化することにより、遺伝子発現を促進する。 一方、JNKはMAPKとは異なり、ストレスやサイトカインなどの刺激、Gタンパク質共役型レセプターのアゴニスト刺激などにより活性化することがわかっている。JNKの上流として、JNK kinase(JNKK)やMEKK,さらにmixed lineage kinase(MLK)などが同定されているが、そのシグナル伝達経路はそれぞれの刺激により異なる。また、JNKも活性化されると核内に移行し、c-Jun及びTCFをリン酸化し活性化することが示されている。従って、JNKがzif268の遺伝子発現に関与する可能性も考えられる。そこで、MAPK及びJNKがNGF或いはカルバコール刺激によるzif268遺伝子の発現に関与するか否かについて調べることにした。 まず、NGF刺激によるzif268遺伝子の発現におけるMAPKの役割を調べるために、薬理学的及び分子生物学的手法を用いて解析を行った。その結果、MEK(MAPKK)阻害剤であるPD098059の前処理によりNGF刺激によるMAPKの活性化を完全に抑制してもzif268遺伝子の発現は部分的にしか抑制されなかった。従って、NGF刺激によるzif268遺伝子の発現はMAPK依存性の経路とMAPK非依存性の経路を介して起こることが示唆される。また、MAPK非依存性の経路はPI3-kinase特異的阻害剤であるwortmanninを低濃度で処理することにより大部分が抑制されるため、MAPK非依存性のzif268遺伝子発現はPI3-kinaseを介することが示唆される。ところが、wortmannin単独で前処理してもNGF刺激によるzif268mRNAの誘導は影響を受けないことから、このwortmannin sensitiveな経路はMAPKが活性化しているときには機能しないことも示唆される。また、PI3-kinaseの下流を特定するため、その候補であるJNKを調べたところ、NGF刺激によるJNKの活性化はPI3-kinase阻害剤で前処理しても抑制されずむしろ増強されることがわかった。従って、NGF刺激によるzif268遺伝子発現において、JNKはPI3-kinaseの下流では機能しないことが示唆される。しかし、JNKの活性を阻害するJNK interacting protein(JIP-1)を細胞内で発現させるとzif268プロモーターの転写活性が部分的に抑制されることから、JNKはzif268遺伝子の発現に関与していることが示唆された。以上の結果より、NGF刺激によるzif268遺伝子発現にはMAPK,JNK,PI3-kinaseのそれぞれが役割を果たすことが示唆される。 次に、カルバコール刺激により、MAPK及びJNKがどのような経路を介して活性化するのかを調べた。mAChRはGタンパク質共役受容体であり、M1からM5までのサブタイプが存在する。M1,M3,及びM5サブタイプは、百日咳毒素(PTX)非感受性のGタンパク質であるGqサブタイプと共役する。一方、M2及びM4サブタイプは、PTX感受性のGタンパク質であるGi/Goサブタイプと共役する。PC12D細胞にはM1及びM4サブタイプが発現しているが、PC12D細胞においてカルバコール刺激によるMAPK及びJNKの活性化は百日咳毒素(PTX)で前処理しても抑制されなかったことより、両者の活性化はM1サブタイプを介すると考えられる。M1レセプターが活性化されるとGqが活性化され、phospholipase C(PLC)が活性化される。活性化したPLCは、細胞膜リン脂質からdiacylglycerol(DAG)及びIP3を産生する。DAGはPKCを活性化し、IP3は細胞内のカルシウムストアからカルシウムを放出させることにより、細胞内のカルシウム濃度の上昇を引き起こす。PKCはRaf-1の活性化を介してMAPKを活性化することが示されている。また、PC12細胞において、細胞内カルシウムの濃度上昇はチロシンキナーゼPYK2及びRasを介してMAPKを活性化することが示されている。そこで、PC12D細胞におけるカルバコール刺激によるMAPKの活性化を調べたところ、dominant-negative RasであるN17Rasを発現させても大部分は抑制されなかった。また、Ras非依存性のMAPKの活性化はPKC阻害剤であるGF109203Xで前処理しても抑制されないが、EGTAの前処理により細胞外からのカルシウムの流入を阻害すると完全に抑制されることがわかった。以上のことから、PC12D細胞におけるカルバコール刺激によるMAPKの活性化は、これまで報告されていた経路とは異なり、主にRasに依存せず、細胞外からのカルシウムの流入に依存する経路を介することが示された。一方、カルバコール刺激によるJNKの活性化はN17Rasの発現により大部分が抑制され、GF109203X或いはEGTAで前処理してもほとんど抑制されないことがわかった。さらに、MEK阻害剤であるPD098059で前処理することによりMAPKの活性化を抑制すると、zif268遺伝子の発現の大部分が抑制されることから、カルバコール刺激によるzif268遺伝子発現の大部分はMAPKを介して起こることが示唆される。 本研究の過程において、zif268遺伝子発現におけるPKCの役割を調べるために従来PKC阻害剤として用いられていたH7[1-(5-isoquinolinesulfonyl)-2-methylpiperazine]を用いて解析を行った。その結果、H7はホルボールエステルによるMAPKの活性化を抑制しないが、zif268及びc-fos遺伝子発現を抑えることがわかった。そこで、H7の細胞内における作用を調べた結果、mRNAの転写伸長に必須であるRNA polymerase IIのリン酸化が阻害されることがわかった。以上の結果から、H7は細胞内においてRNA polymerase IIのリン酸化を阻害することによりzif268及びc-fos遺伝子発現を抑制する可能性が示唆された。 |