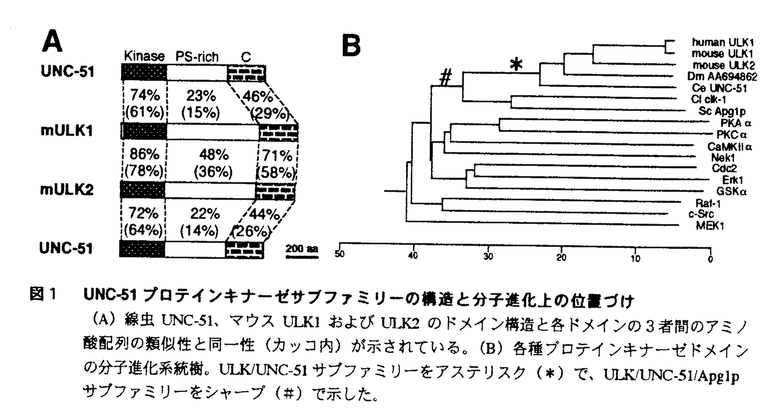
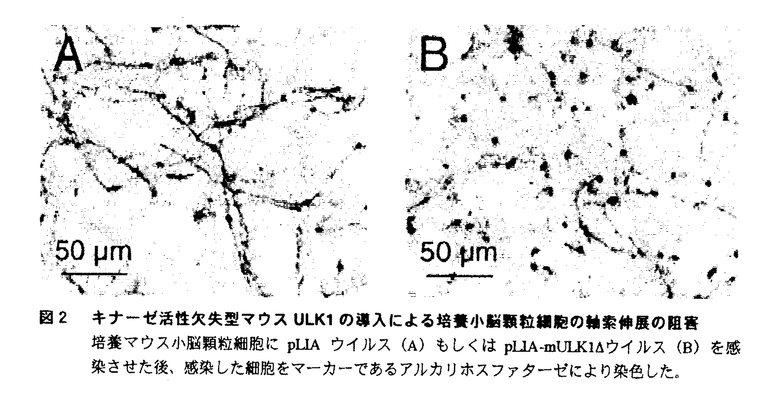
本論文は5章からなる。第1章は本論文全体の序論である。第2章で哺乳類の新規セリン/スレオニンキナーゼULK1およびULK2のcDNAクローニングと基礎的な分子生物学的解析について述べられ、第3章でULK1およびULK2の機能解析について述べられている。第4章は本論文全体の考察であり、第5章に参考文献の一覧が示されている。 論文提出者は、線虫Caenorhabditis elegansで見出されたセリン/スレオニンキナーゼUNC-51が神経細胞の軸索伸展の制御に必須であることに着目し、UNC-51の哺乳類ホモログの単離を試みた。変性プライマーを用いたPCR法によりラット胎仔脳のcDNAからUNC-51に相同性の高いcDNA断片が得られ、この断片をプローブにヒトおよびマウスcDNAライブラリーから、UNC-51と全長にわたって相同性を有する2種類の細胞質型セリン・スレオニンキナーゼのcDNAを単離し、ULK1(UNC-51-like kinase 1)およびULK2と命名した。キナーゼドメインはULK1/ULK2/UNC-51のN末端側に位置し、中央部をPS-richドメイン、C末端領域をC-ドメインと命名した。 アミノ酸配列の相同性解析の結果、ULK1およびULK2はUNC-51の他、酵母Saccharomyces cerevisiaeのオートファジーに必須のApg1p、マメ科の植物に繁殖する菌類Colletotrichum lindemuthianumの感染性に必須のclk-1と相同性が高く、ULK1/ULK2/UNC-51/Apg1p/clk-1は、プロテインキナーゼスーパーファミリーの中で構造上新たなサブファミリーを形成することを示した。 ULK1およびULK2の組織発現分布をノザンブロット法およびin situハイブリダイゼーション法で解析し、ULK1およびULK2が心臓、脳、骨格筋、腎臓、精巣、肝臓、脾臓、肺の各臓器で普遍的に発現していること、マウスの胎仔脳で分化・成熟課程の神経細胞で発現していることを見出した。 ULK1染色体遺伝子の解析を行い、ヒトゲノム上に30〜40kb、マウスゲノム上に24kbにわたって単一コピー遺伝子として存在していることを示した。また、既知の疾患や変異体との関係を探るために、FISH法およびRH法によりULK1遺伝子およびULK2遺伝子の染色体マッピングを行ったが、特定の変異との連関はこれまでのところ見出されていない。 ULK1およびULK2のin vitroでのキナーゼ活性について検討した。ULK1、ULK2はミエリン塩基性タンパク質、カゼイン、ヒストンは基質としないが自己リン酸化能を有することを明らかにした。また、短縮型ULK1、ULK2の解析によって、いずれもPS-richドメインに自己リン酸化を受けることを示した。 マウスULK1のPS-richドメインに対するペプチド抗体を作製し、免疫細胞化学的解析によって、ULK1が細胞質に局在すること、分散培養小脳顆粒細胞の細胞体および神経突起に広く分布することを明らかにした。 続いて、レトロウイルスベクターを用いて培養マウス小脳顆粒細胞にキナーゼ活性欠失型マウスULK1(mULK1 )遺伝子を導入することにより神経突起の伸展が阻害されることを示して、ULK1/ULK2が哺乳類の神経細胞の神経突起伸展においても重要な役割を担っていることを示した。 )遺伝子を導入することにより神経突起の伸展が阻害されることを示して、ULK1/ULK2が哺乳類の神経細胞の神経突起伸展においても重要な役割を担っていることを示した。 さらに、目的遺伝子を線虫unc-51プロモーターで発現可能なベクターを構築し、線虫とマウスの対応する機能ドメインを入れ換えた各種キメラキナーゼを発現するトランスジーンをunc-51変異体に導入した。その結果、ULK1、ULK2のキナーゼドメインおよびPS-richドメインは機能的にも線虫のUNC-51と等価であり、UNC-51の機能にはサブファミリー特異的キナーゼ活性および種特異的C-ドメインの機能が必須であることを示した。 これらの解析から、論文提出者は、ULK1およびULK2がUNC-51とともに構造的にも機能的にも保存された新規のプロテイン・キナーゼサブファミリーを形成することを提唱した。 なお、本論文第2章のULK1およびULK2のcDNAクローニングと塩基配列決定にはヘリックス研究所村松正明博士、in situハイブリダイゼーションによるULK1の発現解析はロックフェラー大学友田利文博士、FISH法およびRH法によるULK1およびULK2遺伝子の染色体マッピングは名古屋大学農学部松田洋一博士、帝京大学医学部高野貴子博士、かずさDNA研究所関直彦博士に技術的支援を受けたが、ULK1およびULK2の遺伝子の分離、発現解析、機能解析など本研究の主要部分は論文提出者が主体となって行ったものであり、論文提出者の寄与が十分であると判断する。 したがって、博士(理学)の学位を授与できると認める。 |