序 リン酸は、窒素、カリウムとともに植物栄養の三大要素として植物体の成長に不可欠であることが古くから知られている。また、リン酸は核酸、リン脂質などの細胞の構成成分であり、代謝経路の中間産物として生命活動に不可欠な成分であることも周知のことである。しかし、植物体内に栄養素として取り込まれたリン酸が、その後どのような生理現象の制御に関わっているのかは明確でない。この点について、酵母のPHO系では培地中のリン酸により制御される、酸性、アルカリ性ホスファターゼ活性の調節機構が示されているが、植物細胞においても酵母リン酸トランスポーター遺伝子のホモログが単離され、リン酸飢餓状態で発現することから、培地中のリン酸の有無と取り込みとの間には調節機構があることが予想される。
このような背景から、植物細胞のリン酸により制御される現象から、リン酸の関与する現象の分子機構が解明されるものとして本研究を開始した。この実験系にはタバコBY-2細胞の培養系を用いることが有効であると考えたが、その根拠として、BY-2細胞は培養細胞の中でもっとも増殖が速く、アフィディコリン、プロピザミドを使って高い同調率が得られることから細胞周期の解析に適した材料であること、また、初期の研究において培地中のリン酸の消費速度が他の無機イオンやショ糖に比べてとりわけ早く、培地中のリン酸の初期濃度と定常期の細胞数とには比例関係が見られることからBY-2細胞の増殖とリン酸との関係が指摘されてきたことが挙げられる。リン酸が細胞増殖の限定要因であることはニチニチソウ培養細胞での予備的な研究から知られていたが、BY-2細胞ではそれがより特徴的であることが、倉屋により示されている(倉屋芳樹、修上論文、1996年)。
本研究では、リン酸を飢餓した細胞にリン酸を再添加することで半同調的な細胞分裂が誘導されることを示し、このリン酸飢餓再添加の系の特徴を同様な細胞分裂を誘導するオーキシン飢餓再添加の系と比較しながら調べた。次に、この系において単離されたリン酸誘導遺伝子phi-1などの発現解析をすることでこの系の特徴の分子的な裏付けをおこない、さらに、リン酸の再添加によって誘導される細胞分裂の機構を解明するために、リン酸誘導遺伝子の単離をおこなった。
結果1.リン酸飢餓再添加による細胞周期の同調 植え継ぎ後8日目のBY-2細胞をリン酸を含まない培地に植え継いだところ、翌日に分裂指数(MI)が約5%に上昇したが、植え継ぎ3日後にはMIは1%以下に低下した。そこへ370mg/lのKH2PO1を添加したところリン酸添加12時間後からMIが上昇し始め、16時間後には26±1.5%に達した(図1)。また、BrdUの核への取り込みによりDNA合成をしている細胞を調べたところ、リン酸添加4時間後からS期の細胞は増え始め、8時間後には18%に達した(図1)。このことから、定常期のBY-2細胞をリン酸を含まない培地で培養すると、リン酸飢餓状態となって細胞分裂が停止し、そこへリン酸を再添加すると、G1期で停止していた細胞周期が再び進行し、DNA合成を伴う半同調的な細胞分裂が誘導されることがわかった。
そこで、より適当な条件を探すためにリン酸飢餓の期間を3日間から7日間、13日間に延長し、リン酸を再添加したところMIの上昇開始時間は遅れ、ピークの値は下がったものの半同調的な細胞分裂の誘導が観察された(図2)。しかし、リン酸飢餓期間を20日間に延長した場合は半同調的な細胞分裂の誘導は観察されなくなった。同様な半同調的な細胞分裂が誘導される系としてオーキシンの飢餓再添加の系が知られているが、この系では3日間の飢餓処理はオーキシンの再添加により半同調的な細胞分裂が誘導されるが、飢餓期間を4日以上に延長すると死んだ細胞が出現しはじめ、半同調的な細胞分裂が観察されなくなる。これより、リン酸の飢餓再添加とオーキシンの飢餓再添加とでは、飢餓状態での細胞の状況および再添加による細胞周期再スタートのしくみが異なると考えられる。
リン酸飢餓細胞とオーキシン飢餓細胞との差異を細胞レベルで探るために、細胞をDAPI染色して観察したところ、オーキシン飢餓細胞と比較して、リン酸飢餓細胞では色素体DNAの蛍光輝度が低くなった(図3A、D)。そこへリン酸を再添加すると4時間後には核の周辺部で、10時間後には細胞質の色素体DNAの輝度が増加した(図3B、C)。これよりリン酸の再添加により色素体DNAの複製が行われるが、それは細胞核DNAの複製に先立っていることがわかった。
なお、細胞をリン酸飢餓条件下で培養すると、通常の培養条件下およびオーキシン飢餓細胞ではほとんど見られない細胞核を2つ持つ細胞が蓄積し(図4A)、6日後には13%に達した。アニリンブルーで細胞を染色し観察したところ、細胞板を構成するカロースの伸長が不十分であった(図4B)。そこで、抗チューブリン抗体染色により微小管構造を観察したところ、フラグモプラストの構造には異常はなかったことから(図4D)、リン酸飢餓条件下では細胞板構成成分の蓄積が不十分になり、正常な細胞質分裂が行われないことが二核の細胞を生じる原因と考えられる。リン酸を再添加するとこれらの二核細胞は不充分な細胞板を保ったまま次の細胞周期に入った(図4E、F)。これより、リン酸は正常な細胞板の形成にも関わっていることがわかった。
これらの結果から、BY-2細胞ではG1期にリン酸の有無により細胞周期の進行を制御する機構が存在し、リン酸はDNA複製の基質として使われる一方で、リン酸飢餓により細胞周期を停止していた細胞を、再び細胞周期に戻す制御因子として働いていると考えられる。しかし、リン酸はオーキシンとは別の経路において細胞周期の再スタートに関わっていると考えられる。
2.リン酸誘導遺伝子phi-1の発現解析 このリン酸飢餓再添加による細胞分裂の誘導のしくみを分子レベルで解明するには、リン酸により発現が制御される遺伝子を単離、解析することが有効である。これまでにディファレンシャルプラークハイブリダイゼーション法により、リン酸により発現が誘導される遺伝子phi(phosphate-induced gene)-1が単離されているが、この遺伝子の全長と思われるcDNAのシークエンスを決定し、発現解析をおこなった。
相同性検索をしたところ、これまでに報告された遺伝子のうちでphi-1と相同性のあるものは見つからなかったが、細胞膜ATPaseのATP結合領域と考えられるLys-Gly-Ala配列が保存されていた。
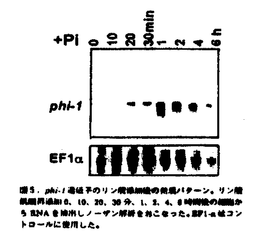
ノーザン解析では、phi-1はリン酸添加20分後から発現が見られ、1、2時間程度で発現量は最大になるが、DNA合成の始まる6時間後には発現は減少したことから(図5)、phi-1遺伝子はリン酸の添加に一過的に応答して発現することがわかった。そこで、アフィディコリン処理により細胞周期を同調化したBY-2細胞におけるphi-1遺伝子の発現量を調べたが、細胞周期の進行に伴う発現量の変化は見られなかったことから(図6)、phi-1遺伝子の発現は細胞周期には依存していないことがわかった。
次に、phi-1遺伝子とオーキシンとの関係を調べるために、phi-1遺伝子の発現をオーキシン飢餓再添加の系で調べたところ、オーキシン飢餓細胞でもオーキシン添加後の細胞でも同様に発現が見られた(図7B)。これより、phi-1はオーキシンによって誘導される細胞分裂とは別の経路において働いていると考えられるが、これは、リン酸とオーキシンとは別の経路で働いているという現象レベルでの考察を裏付けるものである。
そこで、リン酸とオーキシンの両方を飢餓し再添加したところ、リン酸のみの飢餓再添加にくらべて、MIの上昇開始は遅れピークの値も小さくなったが、半同調的な細胞分裂の誘導が観察された。しかし、phi-1はリン酸、オーキシン再添加少なくとも2時間後には発現が見られたことから(図7C)、リン酸、オーキシンの二重飢餓は、phi-1の発現より後の過程にも影響を与えると考えられる。
次に、phi-1遺伝子産物に対する抗体を作製し、遺伝子産物の蓄積を調べたところ、リン酸添加2時間後には遺伝子産物の蓄積が見られ、6時間後でも同様に蓄積していた(図8)。この抗体を使って細胞内の遺伝子産物の分布を調べたところ、リン酸飢餓細胞では遺伝子産物の蓄積は見られなかったが、リン酸添加6時間後の細胞では核の周辺部、および、細胞表層に網目状の染色が見られた(図9)。
タバコ植物個体の各器官におけるphi-1の発現を調べたところ、poly(A)+RNAを使ったノーザン解析では花、茎、葉、根、いずれの器官で転写産物を確認できたが、全RNAを使った場合ではほとんどシグナルは検出されなかった(図10)。これより、植物体の通常の成育状態ではphi-1遺伝子産物の蓄積は少ないと考えられる。
これらの結果から、phi-1遺伝子はリン酸飢餓により細胞周期の進行が停止している細胞においてリン酸に対し急速に応答し、リン酸の再添加によって生じる色素体DNA、細胞核DNAの複製に先立って発現することがわかった。しかし、phi-1遺伝子の発現は細胞周期に依存しないことから、phi-1は細胞周期の制御因子としてではなく、G1期で細胞周期を停止していた細胞を再び細胞周期にを戻すことに働いていると考えられるが、オーキシンによる細胞周期の進行とは別の経路で働いていることがわかった。phi-1遺伝子産物には、細胞膜ATPaseのATP結合領域と考えられるLys-Gly-Ala配列が保存されていることから、phi-1遺伝子産物は何らかのタンパク質のリン酸化を通して働いている可能性がある。
3.リン酸により発現が誘導される遺伝子の探索 このリン酸飢餓再添加の系には、phi-1遺伝子以外にもリン酸によって発現が制御される遺伝子があると考え、より感度の高いディファレンシャルディスプレイ法をもちいてさらにスクリーニングをおこなった。その結果、新たに2つのリン酸誘導遺伝子を単離し、それぞれphi-2、phi-3と名付け、解析をおこなった。
phi-2遺伝子はC端側にbZIP型転写因子と高い相同性を持つ領域を含んでいたが、全長にわたって高い相同性を持つ遺伝子は見つからなかったことから、phi-2は新規のbZIP型転写因子をコードしていると考えられる。
ノーザン解析では、phi-2遺伝子は発現量が少ないため、poly(A)+RNAをもちいた。リン酸飢餓細胞に比べてリン酸添加2時間後の細胞では転写産物量は増えたが、4、6時間後の細胞では減少したことから(図11)、phi-2遺伝子はリン酸の再添加によって生じる色素体DNAの複製よりも早い時期に一過的に発現することがわかった。
いっぽう、phi-3遺伝子は真核生物のタンパク質翻訳開始因子(eIF)-5と高い相同性が見られ、eIF-5に特徴的であるGTPaseモチーフ(GXGNGIKT)、およびrRNAと結合すると考えられるジンクフィンガーを形成する4個のCys残基が保存されていたことから、phi-3はeIF-5をコードしていると考えられる。
phi-3遺伝子の発現量はphi-2遺伝子と同程度で少ないが、リン酸添加2時間後には転写産物の蓄積が見られ、6時間後には発現が減少した(図12)。
これら、phi-2、phi-3遺伝子の発現のタイミングをphi-1遺伝子と比較すると、リン酸添加20分後という比較的早く発現するphi-1に対し、phi-2は1、2時間後、phi-3は2、4時間後という一段階遅れた発現をする。リン酸飢餓細胞ではリン酸の再添加によりヌクレオチドなどのリン酸を含む細胞構成成分の再合成が必要であり、この時期に発現するphi遺伝子群はこの過程に関わっていると考えられる。まず、リン酸の再添加により、転写、翻訳因子をコードすると考えられるphi-2、phi-3が発現することから、細胞内での転写、翻訳レベルが活性化される。そして、これらの新規の転写、翻訳装置によって合成された構造タンパク質により、細胞構成成分の再合成がおこなわれるという調節機構が予測される。リン酸は細胞構成成分の素材であるが、それは単に素材として利用されるのではなく、リン酸自身を出発点とした制御機構によって、再合成の時期や量の調節を受けながら細胞構成成分に取り込まれ、さらに、これらの成分の再合成が細胞周期進行のシグナルとして働いていると考えられる。



 m。
m。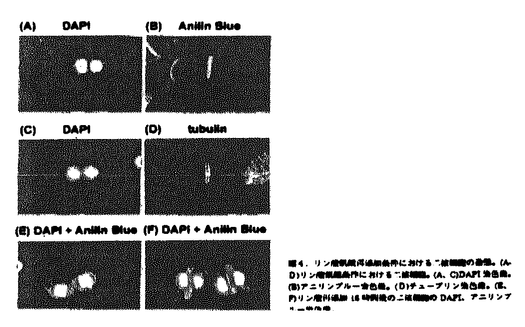
 はコントロールに使用した。
はコントロールに使用した。





