生物はその種内に多くの遺伝的変異(多様性)を持っていることが知られている。たとえば、エンドウマメの皺の有無や人間のABO血液型などが種内変異の典型的な例であり、それぞれが対立遺伝子といわれている。集団遺伝学では、どのようなメカニズムによってこのような種内変異が維持されているのかを研究することにより、生物の進化機構を考える。現在観察された種内変異の量及びパターンを説明するのに自然選択の力が必要なのかどうかが、集団遺伝学の議論の中心になっている(自然選択仮説と中立仮説)。そのため、我々がある種内変異を観察したとき、その変異の生まれた要因や、現在に至るまでの過程を考える必要がある。このとき、確率過程をもとにした数学理論が非常に役に立つ。 本研究では、集団中に対立遺伝子が2つ存在するとき、その遺伝子座領域におけるDNAレベルの変異が、どのようなかたちで観察されるかを理論的に解析した。2つの対立遺伝子クラス(allelic class)をそれぞれA1、A2と表す。そして、n個の個体をサンプルしたとき、2つの対立遺伝子クラス、A1及びA2、の個数がそれぞれiとn-iであったとする。このときの対立遺伝子クラス内及びクラス間の変異量について、理論的な研究をおこなった。一般的に種内の変異量は、2つのパラメーター(S:No.of segreagting sites、K:Average no.of pairwise differenes)で表される。A1対立遺伝子クラス内の変異量(S1(i,n-i),K1(i,n-i))、A2対立遺伝子クラス内の変異量(S2(i,n-i),K2(i,n-i))、及び両対立遺伝子クラス間の変異量(Sb(i,n-i),D(i,n-i))の中立仮説のもとでの期待値を、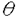 の関数で表すことができた。ここで、(=4N の関数で表すことができた。ここで、(=4N )は集団中の変異量を表すパラメーターである(Nは集団の大きさ、は突然変異率)。すなはち、No.of segreagting sitesは、 )は集団中の変異量を表すパラメーターである(Nは集団の大きさ、は突然変異率)。すなはち、No.of segreagting sitesは、    ここで  また、Average no.of pairwise differenesは、 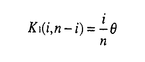  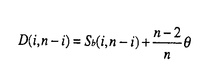 FIGURE1は、n=20のときの数値解をプロットしたものである。また、FIGURE2では、A1対立遺伝子クラスが祖先型でA2対立遺伝子クラスが突然変異型であるという条件のもとでの数値解をプロットした。 次に、2つの対立遺伝子クラスが自然選択の力によって維持されているときの、対立遺伝子クラス内及びクラス間の変異量について研究した。もちいた自然選択モデルは、genic selection modelとoverdominant selection modelの2つである。この2つの自然選択モデルのもとで、対立遺伝子クラス間の変異量(D(i,n-i))の期待値を理論的に求めることができた。FIGURE3は、n=10のときの数値解をプロットしたものである。対立遺伝子クラス内の変異量(K1(i,n-i)とK2(i,n-i))に関しては、コンピューターシミュレーションを用いて調べた。FIGURE4に、その結果を示す。 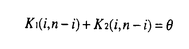 という中立仮説のもとで成り立つ関係が、モデル及び自然選択の強さに関わらず、近似的に成り立つことがわかる。 これらの理論的結果が、実際の種内変異のデータを理解するにあたって、どのように役に立つかを考える。  FIGURE1-The amounts of nucleotide variation within and between allelic classes when the ancestral class is unknown FIGURE1-The amounts of nucleotide variation within and between allelic classes when the ancestral class is unknown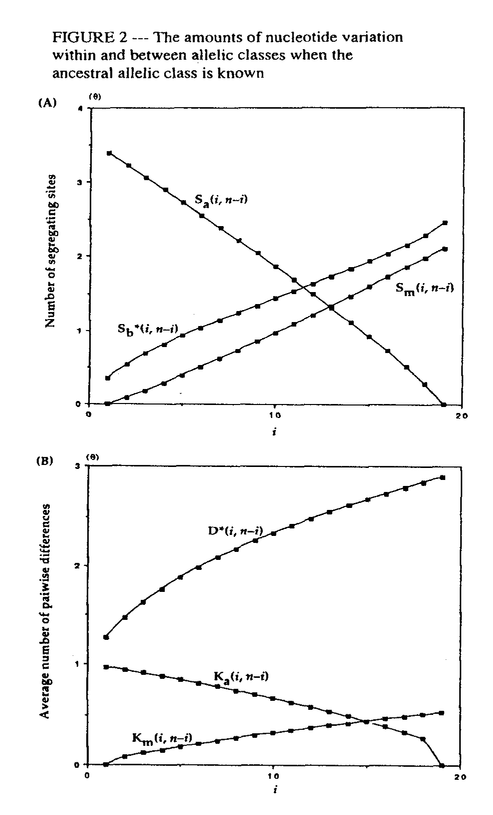 FIGURE2-The amounts of nucleotide variation within and between allelic classes when the ancestral allelic class is known FIGURE2-The amounts of nucleotide variation within and between allelic classes when the ancestral allelic class is known FIGURE3-Average number of pairwise differences between two allelic classes under selection model FIGURE3-Average number of pairwise differences between two allelic classes under selection model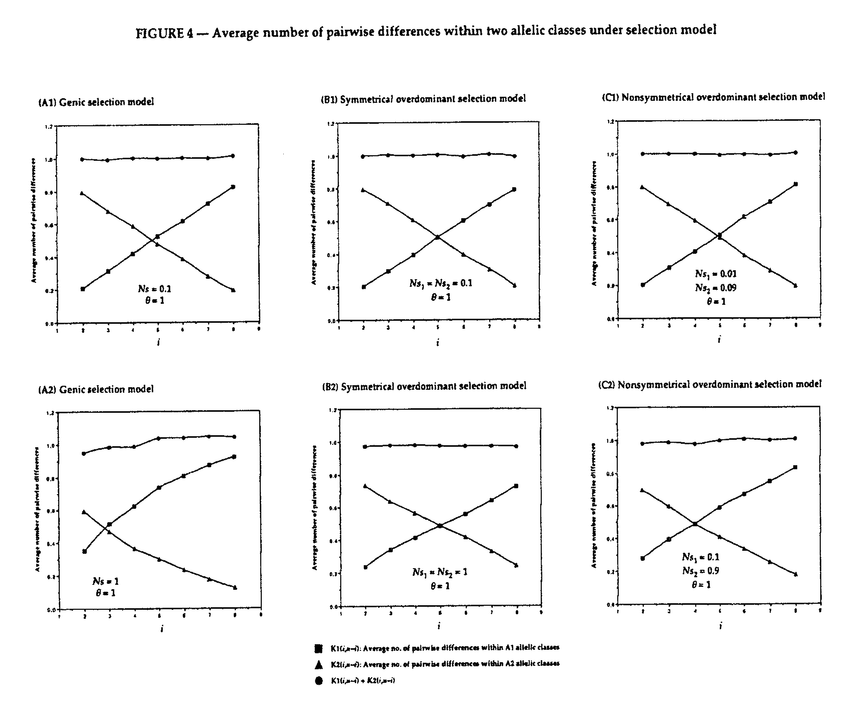 FIGURE4-Average number of pairwise differences within two allelic classes under selection model FIGURE4-Average number of pairwise differences within two allelic classes under selection model |