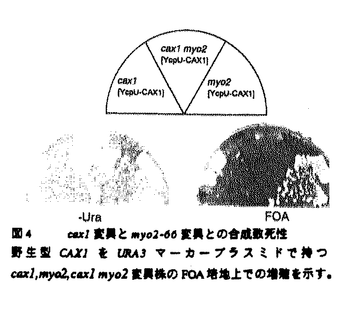
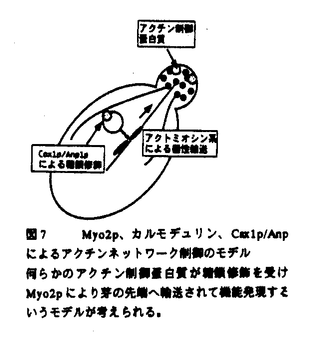
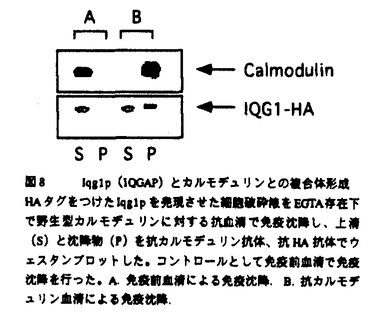
本論文は3章からなり、第一章はカルモデュリンのターゲット蛋白質であるMyo2pとの相互作用の解析、第二章はcmd1-226と合成致死性を示す変異株の解析、第三章では、新規カルモデュリン結合蛋白質を解析した。 まず、第一章ではアクチン結合部位に変異を持つmyo2変異株(myo2-66)とcmd1-226の細胞形態が類似していること、myo2-66変異がcmd1-226とアレル特異的な合成致死を示すこと、およびCmd1-226pのMyo2pへの結合が他の変異カルモデュリンに比べて特異的に低下していることを明らかにした。これらの結果からMyo2pがカルモデュリンによるアクチンネットワーク制御における標的蛋白質であり、出芽、カルモデュリンの芽の部分への局在には別の標的蛋白質が関与すると予想された。そこでMyo2pがアクチンネットワーク以外のプロセスに関与する可能性を排除するため、myo2-66変異と各機能欠損を示すカルモデュリン変異との2重変異株の増殖を更に詳しく調べた。その結果、myo2-66は、アクチンネットワーク欠損を示すカルモデュリン変異、cmd1-226と合成致死性を示すが、一方でcmd1-228(カルモデュリンの芽の部分の局在に欠損をもつ)、cmd1-233(出芽に欠損をもつ)、cmd1-239 (核分裂に欠損をもつ)とは遺伝的相互作用を示さなかった。これらの結果は、アクチンネットワーク制御におけるカルモデュリンのターゲットはMyo2pでありカルモデュリンの芽の部分への局在、出芽、核分裂のプロセスにおいてはMyo2p以外のターゲットが関与していることを支持している。myo2-66 cmd1-233二重変異株は単独変異株に比べて増殖速度が遅くなっていたが、これはcmd1-233もMyo2pとの相互作用にある程度の欠損を持つことで説明できる。次にmyo2-66とcmd1-226が合成致死になる分子メカニズムを考察した。Cmd1pはMyo2pの正常な局在に必要であるか・またはCmd1pはMyo2pのモーター活性を調節していると考えられたので、cmd1-226変異様におけるMyo2pの局在を調べた。野生株ではMyo2pは小さな芽をもつ細胞の先端に存在するが、cmd1-226変異株においても野生株と同様のMyo2pの局在が観察された。ミオシンIIファミリーの生化学的研究から、ミオシン軽鎖を持たないミオシンのATPase活性・およびモーター活性が低下することが知られている。従って、カルモデュリンはMyo2pの局在ではなく、活性を制御すると考えている。またアクチンの局在性が失われていてもMyo2pの芽の部分の局在は正常であることから、Myo2pはカルモデュリンにもアクチンにも依存しない機構で芽の先端部分に局在していることが示唆されれた。カルモデュリン結合ドメインを欠失したMyo2pが野生株と変わらず芽の先端に存在するという報告(Stevens and Davis1998)、および、アクチン重合阻害剤を加えた細胞においてもMyo2pは芽の先端部分に存在する(Ayscough et al.,1997)という報告もこの考えを支持している。 第2章では、カルモデュリン変異と合成致死を示すcax変異株の遺伝的相互作用を調べた。その結果、cax1変異のみがmyo2-66変異と同様、cmd1-226とアレル特異的に合成致死になることを見いだした。cax1変異はmyo2-66変異とも合成致死性を示す。この性質からCAX1遺伝子がカルモデュリン・Myo2pによるアクチンネットワーク制御に特に深い関わりを持つことが考えられる。またcax2,cax3変異はcmd1-226,cmd1-228,cmd1-233と、cax4変異はcmd1-226,cmd1-228と、cax5変異はcmd1-226,cmd1-233と合成致死を示した。cax2,cax3,cax4,cax5が細胞極性に欠損を示すカルモデュリン変異と特に合成致死性を示すことから、Cax2p,Cax3p,Cax4p,Cax5pは広く細胞極性に関与していると考えられる。このうちCAX1,CAX4,CAX5のクローニングを行った。その結果、CAX1はANP1と同一であり、CAX5はMNN10と同一であった。Anp1pもMm10pもゴルジ体での蛋白質への糖外銀の付加に機能を果たしている。CAX4は機能未知のCWH8と同一で、239アミノ酸からなる推定3つの膜貫通領域をもつ蛋白質をコードしている。Myo2pは芽の先端への蛋白質輸送に関与すると考えられていることをふまえ、カルモデュリン、Myo2p,Cax1p/Anp1pによるアクチンネットワーク制御機構を考察した。その際まず1)Myo2pが糖鎖修飾の起こる小胞体->ゴルジ体の分泌過程に必要か、2)Myo2pが糖鎖修飾そのものに必要か、の2点について調べた。まず、cmd1-226変異様において小胞体->ゴルジ体->細胞外の順に輸送される蛋白質、インベルターゼの細胞外への分泌を調べたところ、野生株と同程度であった。また、myo2-66,cmd1-226変異様において輸送蛋白質の糖鎖修飾をインベルターゼを指標に調べたところ、野生株と同様に糖鎖修飾による高分子量へのシフトが見られた。これらの結果はMyo2pが糖鎖修飾の起こる分泌過程にも、糖鎖修飾そのものにも関与しないことを示唆している。そこで、何らかのアクチン制御蛋白質がAnp1pによる糖鎖修飾を受けた後、Myo2pにより芽の先端に運ばれて機能するモデルが考えられる。 カルモデュリンの芽の部分への局在、出芽、という形態形成の過程に関与する標的蛋白質は現在のところ不明である。そこで第三章では、酵母のゲノムデータベースからカルモデュリン結合部位を持つ蛋白質を探索した。その結果、アクチン細胞骨格系の制御・細胞極性に機能を持つことが報告されている(酵母におけるIQGAPホモログ)がカルモデュリンと物理的に相互作用することがわかったIqg1pはIQモチーフを8箇所に持つ蛋白質である。まず、免疫沈降法により、Iqg1pとカルモデュリンがin vivoで複合体を形成することを確かめた。次にゲルオーバーレーアッセイを行ったところ、Iqg1pのIQモチーフとカルモデュリンが直接相互作用することが明らかになった。IQG1は生育に必須な遺伝子であることからも、カルモデュリンの標的蛋白質の有力な候補であると考えられる。 なお、本論文第一章/第二章は、大矢禎一と、David Botstein博士、と共同研究であるが、論文提出者が主体となって分析及び検証を行なったもので、論文提出者の寄与が十分であると判断する。 よって、博士(理学)の学位を授与できると認める。 |