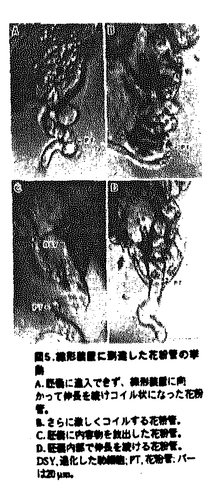
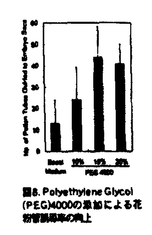
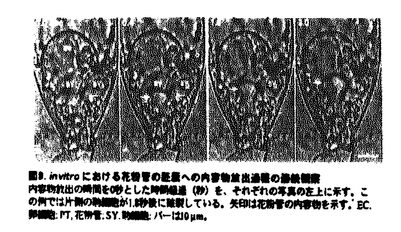
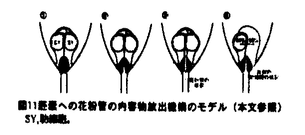
本論文は3章からなり、第1章は、トレニア(Torenia fournieri)の裸出胚嚢へのin vitro花粉管誘導系の確立と花粉管誘導機構の解析、第2章は、in vitro系を用いた花粉管の胚嚢への内容物放出機構の解析、第3章は、重複受精過程における2つの精細胞の動態解析について述べられている。 ゴマノハグサ科のトレニアは卵細胞や中心細胞からなる胚嚢が胚珠組織から突出する高等植物である。本論文では、この裸出胚嚢を持つトレニアの胚珠を花粉管と共培養することにより重複受精をin vitroで再現し、重複受精過程を生体試料を用いて解析している。従来、多くの高等植物では胚嚢が胚珠組織の中央に埋め込まれているために、主に重複受精過程は固定した試料の切片で観察されてきた。一方、in vitroでの受精と言えば、近年は主に酵素処理によって単離された卵細胞や中心細胞が用いられてきた。トレニアの裸出胚嚢に着目し、in vivo同様、胚嚢と花粉管による重複受精をin vitroで再現しようと考えたことは、極めて独創性が高いと評価できる。 本論文は、重複受精過程の進行順に従って解析された内容から構成されている。第1章においては裸出胚嚢への花粉管誘導系を確立している。これまでに多くの植物で胚珠と花粉管を用いたin vitro受精系が確立されているが、このトレニアの系は胚嚢と花粉管の直接的な相互作用を解析できる点で全く新規の系である。剥き出しの胚嚢への花粉管誘導は非常に困難な試みであったが、花粉管と胚嚢の両方に適した培地の開発、超低融点アガロースで固化した培地での胚珠の培養、花柱組織を通過させた花粉管を用いるなどの様々な工夫で、花粉管を直接胚嚢へ誘導することに成功している。この系の確立によって、花粉管は胚嚢の周りに伝うための組織がなくても、正確に胚嚢へ誘導されることが証明された。さらに、胚嚢への誘導活性が、未受精かつ生きた胚嚢の線形装置領域から伝播し、若干の種特異性を有することが示された他、誘導される花粉管の反応性が雌蕊組織を通過することによって向上することなどの多くの重要な知見が得られた。 第2章においては、第1章で確立した系をさらに改良し、花粉管が胚嚢へ内容物を放出する過程について解析している。これにより、花粉管が胚嚢へ内容物を放出する瞬間の映像を、世界で初めて撮影した。この映像を解析することにより、花粉管の先端が胚嚢内部の細胞間隙で破裂し、引き続いて平均わずか0.9秒後に2つの助細胞の片側だけが選択的に崩壊し、もともと助細胞が占めていた領域が花粉管と助細胞の内容物で満たされることが明らかになった。これは、従来予想されていた花粉管が助細胞の内部に進入して内容物を放出するという説とは、異なるものである。花粉管が破裂する正確な場所を特定したことは、今後花粉管の破裂を引き起こすメカニズムや助細胞崩壊のメカニズムを明らかにしていく上で、極めて重要な発見である。 さらに第3章では、SYTOX Greenを用いた精細胞の蛍光生体染色法、ならびに高感度冷却CCDカメラを用いた超微量蛍光観察法を確立し、重複受精過程における2つの精細胞の動態を可視化することに成功している。特に、胚嚢内部でそれぞれの精細胞が動く様子が捉えられたことから、鞭毛を持たない高等植物の精細胞が、胚嚢内部を移動して確実に受精する未知の機構が存在することが示唆された。さらに、花粉管が内容物を放出した胚嚢において、in vivo同様の初期胚発生および初期胚乳形成が観察された。これは、in vivoを反映したin vitro系の確立に成功したことを物語っている。 本論文で開発されたin vitro重複受精系は、花粉管誘導物質や花粉管の反応性を向上させる雌蕊側因子の同定に、花粉管誘導のモデル系として利用できる。また、重複受精を操作した上で完全種子を作り出す系として利用することなども可能であり、この系の重要性や、応用性は極めて高いと言える。トレニアは、安定した形質転換も可能な植物であることから、分子生物学的な解析への展開も期待される。以上のことから、本論文は極めて独創的かつ斬新な論文であるのみならず、今後の高等植物の生殖研究の一大分野の礎を築いた重要な論文であると結論できる。 なお、本論文第1章は、黒岩晴子、河野重行、黒岩常祥の共同研究であるが、論文提出者が主体となって分析及び検証を行なったもので、論文提出者の寄与が十分であると判断する。 したがって、博士(理学)の学位を授与できると認める。 |  m/secに近い順方向および逆方向の激しい原形質流動の中で、精細胞は等速度ではなく、速度を変えながら、止まったり、後退したりしながら、しだいに花粉管の伸長方向に向かって移動することがわかった(図13、図14)。精細胞と栄養核の挙動は連動していることが多いが、しばしば異なる挙動を示し、精細胞と栄養核の距離もダイナミックに変化することがわかった。花粉管の内部では2つの精細胞は常に対になって移動した。さらに、胚嚢に放出された2つの精細胞が、それぞれ別個に動く様子を観察すること成功した(図15)。被子植物の精細胞は鞭毛を持たないが、何らかの機構で移動し、選択的かつ確実に受精すると考えられる。
m/secに近い順方向および逆方向の激しい原形質流動の中で、精細胞は等速度ではなく、速度を変えながら、止まったり、後退したりしながら、しだいに花粉管の伸長方向に向かって移動することがわかった(図13、図14)。精細胞と栄養核の挙動は連動していることが多いが、しばしば異なる挙動を示し、精細胞と栄養核の距離もダイナミックに変化することがわかった。花粉管の内部では2つの精細胞は常に対になって移動した。さらに、胚嚢に放出された2つの精細胞が、それぞれ別個に動く様子を観察すること成功した(図15)。被子植物の精細胞は鞭毛を持たないが、何らかの機構で移動し、選択的かつ確実に受精すると考えられる。