リアルタイムに入力される感覚信号の中から、脳はどのようにして重要な情報を抽出し、記憶しているのであろうか。海馬は日常的なイベントを記憶するエピソード記憶に中心的な役割をしていると言われている。しかし、その情報処理のメカニズムについては、まだ明確には分かっていない。本研究では、海馬の記憶のダイナミックな情報処理機能を明らかにするために、海馬と同様の生理学的、解剖学的特性を持つニューラルネットワークモデルを構築し、その動的特性を明らかにすることを目的とする。今まで提案されてきた海馬モデルの多くは、平均発火率の高い細胞の空間パターンを情報表現として仮定してきた。低頻度のスパイクは、コーディングに参加しないノイズとして無視される。しかし、実際の海馬の細胞発火の時空間パターンは、ダイナミックに変化しており、例えばラット海馬の同じ場所細胞が、環境の変化によって全く異なる場所に反応するようになる。海馬は、解剖学的にはdivergent/convergentな結合を持ち、嗅内野の深層から出発して嗅内野の浅層に帰る閉ループネットワーク構造を持つ(図1)。従って、divergent/convergentな結合上を伝わるスパイクと、スパイクパターンに応じて変化するシナプス荷重との相互作用によってダイナミックに動作するシステムと考えられる。その動的特性によって海馬の機能が発現している可能性がある。本論文では、海馬の解剖学的、生理学的データに基づいて構築した、学習メカニズムとスパイクニューラルネットワークモデルについて述べる。また、これらの動的特性と、それから示唆される海馬の情報処理について論じる。  図1:皮質-海馬系の解剖学的構造. 図1:皮質-海馬系の解剖学的構造. 最初に、海馬と同様のdivergent/convergentな閉ループネットワーク構造において、高頻度発火するパスを自己組織化する学習メカニズムを示す。各シナプスに割り当てられたsij(t)の値がある閾値以上になればLTP(長期増強)を起こし、ある閾値以下になればLTD(長期抑制)を起こす。Sijは、次の(1)式に従う。  細胞#jから細胞#iへのシナプス入力がある場合、細胞#iが細胞が発火した時は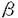 ijは1の値を取り、それ以外の時は-1を取る。従って、Sij(t)の値は入力と発火の相関が高いほど増加し、相関が低いほど減少する。この学習メカニズムでは、局所的ダイナミクスに加え、Fitzsimondsらの海馬細胞における実験データに基づくLTDの逆伝搬という大局性を持つ。これによって、海馬閉ループにおける発火スパイクの相関に応じたシナプス結合パターンが構成される。図2は、以上の学習メカニズムの動作を確認するためのdivergent/convergent結合するニューラルネットワークモデルの構造である。このモデルは、A、B、Cの3層構造から構成され、A層からB層とB層からC層へは投射領域のより広がるダイバージェントな結合を持つ。海馬でC層からA層へとA層からC層へにあたるECからCA1、CA1から海馬台を経由してECへ入る投射は、解剖学的にダイバージェント性は少なく、限定された投射領域を持つ。yiを細胞の出力値、wijをシナプス荷重、Iiを外部入力とすると、各細胞の内部活動度xiは次の式に従う。 ijは1の値を取り、それ以外の時は-1を取る。従って、Sij(t)の値は入力と発火の相関が高いほど増加し、相関が低いほど減少する。この学習メカニズムでは、局所的ダイナミクスに加え、Fitzsimondsらの海馬細胞における実験データに基づくLTDの逆伝搬という大局性を持つ。これによって、海馬閉ループにおける発火スパイクの相関に応じたシナプス結合パターンが構成される。図2は、以上の学習メカニズムの動作を確認するためのdivergent/convergent結合するニューラルネットワークモデルの構造である。このモデルは、A、B、Cの3層構造から構成され、A層からB層とB層からC層へは投射領域のより広がるダイバージェントな結合を持つ。海馬でC層からA層へとA層からC層へにあたるECからCA1、CA1から海馬台を経由してECへ入る投射は、解剖学的にダイバージェント性は少なく、限定された投射領域を持つ。yiを細胞の出力値、wijをシナプス荷重、Iiを外部入力とすると、各細胞の内部活動度xiは次の式に従う。   高頻度に発火する空間パターンが記憶され、想起されることが、コンピュータシミュレーションによって示される(図3)。これは、学習によってシナプス荷重にLTP/LTDが起こり、高頻度発火の入力パターンに応じたシナプス結合パスが自己組織化される(図4)からである。 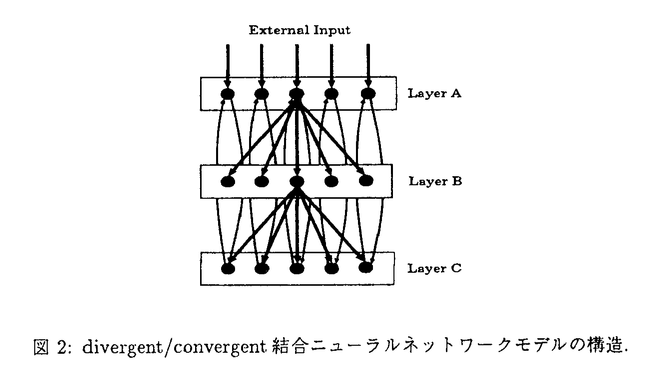 図2:divergent/convergent結合ニューラルネットワークモデルの構造. 図2:divergent/convergent結合ニューラルネットワークモデルの構造. 図3:記憶と想起のシミュレーション(キュー信号による想起). 図3:記憶と想起のシミュレーション(キュー信号による想起). 図4:学習メカニズムによるシナプス荷重の変化(t=300). 図4:学習メカニズムによるシナプス荷重の変化(t=300). 海馬の神経回路網は、大きく階層型ネットワークモデルと相互結合型ネットワークモデル(CA3)とに分けられる。次に、それぞれのモデルの動的特性を示す。図5は、3層(A,B,C)のdivergent/convergentニューラルネットワークモデルに、ポアソン過程によって生成した非同期パターンを入力した時の、外部入力パターンとC層のラスタープロットである。C層で同期的な信号が出現していることが分かる(例えばt=600付近)。図5の外部入力パターンの一部(428 t447)を入力した時のA層、B層、C層の出力をプロットしたのが図6である。t=447でA層の細胞の6個に同期発火が起こり、それがB層、C層への同期発火を引き起こしている。矢印は、同期発生直前の外部入力のA層細胞への入力を示した。t=430あたりから、外部入力がA層の同期発火細胞へ入力されている。t=440からの短い時間内に多くの入力を受けて、同期が起こっている。このように、膜電位の時定数程度の数十msオーダーのタイムウィンドウの時空間パターンが同期発生の原因として働いている。一方、その7、8倍程度の長さのタイムウィンドウ内の微小な変動によって、同期が起こらなくことがある。従って、100ms程度の時空間パターンからある構造が検出されたことを、同期信号が示していると考えられる。また、結合の伝達遅延の分散が小さい場合や部分結合の割合が大きいネットワークほど同期が起こりやすい特性がある。 t447)を入力した時のA層、B層、C層の出力をプロットしたのが図6である。t=447でA層の細胞の6個に同期発火が起こり、それがB層、C層への同期発火を引き起こしている。矢印は、同期発生直前の外部入力のA層細胞への入力を示した。t=430あたりから、外部入力がA層の同期発火細胞へ入力されている。t=440からの短い時間内に多くの入力を受けて、同期が起こっている。このように、膜電位の時定数程度の数十msオーダーのタイムウィンドウの時空間パターンが同期発生の原因として働いている。一方、その7、8倍程度の長さのタイムウィンドウ内の微小な変動によって、同期が起こらなくことがある。従って、100ms程度の時空間パターンからある構造が検出されたことを、同期信号が示していると考えられる。また、結合の伝達遅延の分散が小さい場合や部分結合の割合が大きいネットワークほど同期が起こりやすい特性がある。  図5:階層型ネットワークにおける発現的同期パターン. 図5:階層型ネットワークにおける発現的同期パターン. 図6:同期的発火細胞(A層)への外部入力. 図6:同期的発火細胞(A層)への外部入力. 次に、相互結合をしているスパイクニューロンモデルの発火パターンが、不安定な時空間パターンと安定した平均発火率という2つの特性を持つことを示す。モデルの結合は全結合ではなく、部分的(シミュレーションでは40%)に結合する。yiを内部活動度、xiを細胞の出力値とすると、細胞のダイナミクスは次の(4)式に従う。(4)式の右辺3項目は不応性である。   ポアソン過程系列と、(4)式のAiへの一定値の入力を与えた時のそれぞれのスパイク列は、いずれの入力パターンの場合も非周期的発火パターンが観測される(図8(a)左)。微小摂動を与えた時の軌道からのずれの比の対数の平均は、図7のようになる。パラメータ actを変化させてもほぼ0以上であり、摂動に対して不安定である。同時に、細胞ごとの平均発火率の空間パターン(図8(a)右)は、ニューラルネットワークモデルが同一の場合に安定する。図8(b)は、同一のニューラルネットワークモデルにおいてポアソン過程による入力スパイクの平均スパイク間隔とランダム種を変化させた時の、平均発火率の空間パターンの相関係数である。ネットワークを変化させると相関が低くなるので、図8(b)の高い相関は、平均発火率の空間パターンが入力の時空間パターンに依らず、ネットワーク固有のものであることを示唆する。ニューロンの膜時定数を小さくすると、平均発火率の空間パターンが安定しなくなるので、ニューロンのintegrate-and-fireのためにシナプス結合の構造が現れていると考えられる。これらの結果は、脳内の情報表現において、時空間パターンと平均発火率の空間パターンという2つの可能性を示す。 actを変化させてもほぼ0以上であり、摂動に対して不安定である。同時に、細胞ごとの平均発火率の空間パターン(図8(a)右)は、ニューラルネットワークモデルが同一の場合に安定する。図8(b)は、同一のニューラルネットワークモデルにおいてポアソン過程による入力スパイクの平均スパイク間隔とランダム種を変化させた時の、平均発火率の空間パターンの相関係数である。ネットワークを変化させると相関が低くなるので、図8(b)の高い相関は、平均発火率の空間パターンが入力の時空間パターンに依らず、ネットワーク固有のものであることを示唆する。ニューロンの膜時定数を小さくすると、平均発火率の空間パターンが安定しなくなるので、ニューロンのintegrate-and-fireのためにシナプス結合の構造が現れていると考えられる。これらの結果は、脳内の情報表現において、時空間パターンと平均発火率の空間パターンという2つの可能性を示す。  図7:相互結合型ネットワークにおける摂動の影響 図7:相互結合型ネットワークにおける摂動の影響 図8:相互結合型ネットワークのシミュレーション結果 図8:相互結合型ネットワークのシミュレーション結果 最後に、階層型と相互結合型で構成された海馬全体のニューラルネットワークモデル構成し、前述の学習メカニズムを適用した時のスパイクの挙動を示す。そして、海馬の機能との関連を論じる。モデルの構造は、A、B、Cの3層構造であり、B層だけが相互結合型ネットワークである(図9)。解剖学的なデータを基に、層間の結合割合をA層からB層へが0.25、B層からC層へが0.4、A層からC層へが0.1、C層からA層へが0.02、B層内の相互結合が0.4と設定する。ポアソン過程による系列(ISI=100)を各細胞に入力した時の外部入力のパターンと各層のラスタープロットを、図10に示す。学習メカニズムが働くことによって、A層に周期的な時空間パターンが出力されてくる。これは、B層内のリカレント結合や他の層間で起こったLTDとLTPが効いていると考えられる(図11、図12)。特に、LTDによって相関の少ないスパイクが抑制され、入力の周期性に合うスパイクが全体の閉ループに残ったと考えられる。この結果は、不安定で非周期的なスパイクの挙動が、学習メカニズムによって安定なパターンへ変換される可能性を示唆している。  図9:海馬全体モデルの構造 図9:海馬全体モデルの構造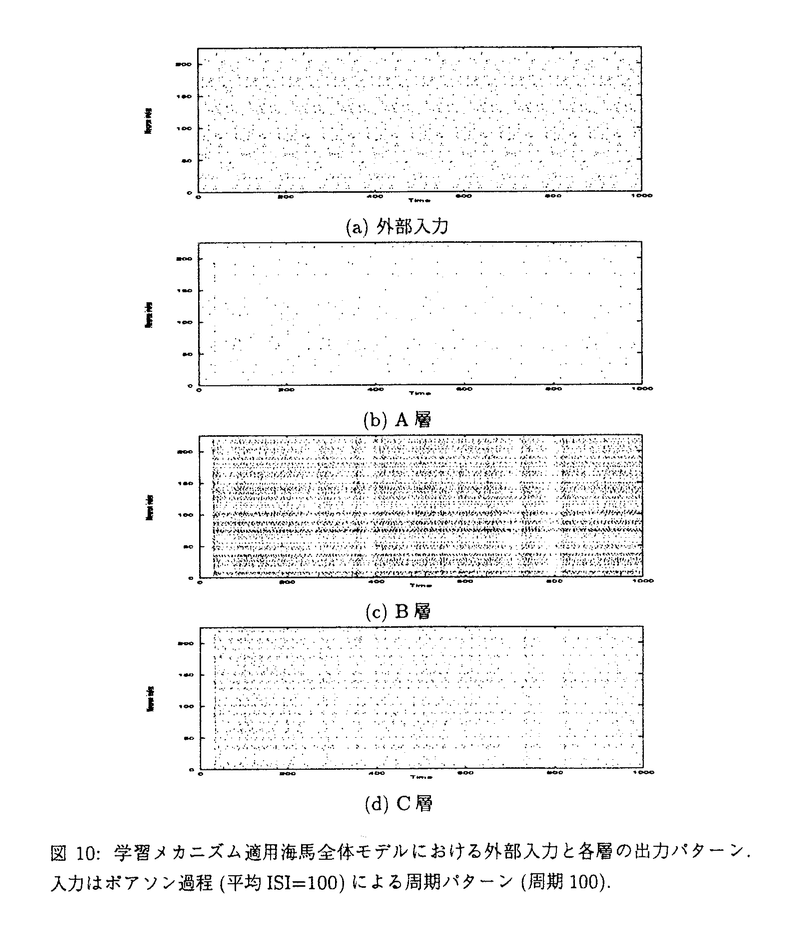 図10:学習メカニズム適用海馬全体モデルにおける外部入力と各層の出力パターン.入力はポアソン過程(平均ISI=100)による周期パターン(周期100). 図10:学習メカニズム適用海馬全体モデルにおける外部入力と各層の出力パターン.入力はポアソン過程(平均ISI=100)による周期パターン(周期100). 図11:LTDの強く起こった結合.左:A層⇒B層,B層⇒C層,C層⇒A層.右:B層内(LTD率<0.4).縦軸はニューロンインデックス. 図11:LTDの強く起こった結合.左:A層⇒B層,B層⇒C層,C層⇒A層.右:B層内(LTD率<0.4).縦軸はニューロンインデックス. 図12:LTPの強く起こった結合.左:A層⇒B層,B層⇒C層,C層⇒A層.右:B層内(LTP率>1.5).縦軸はニューロンインデックス. 図12:LTPの強く起こった結合.左:A層⇒B層,B層⇒C層,C層⇒A層.右:B層内(LTP率>1.5).縦軸はニューロンインデックス. 学習されていないパターンが入力された時、CA3内と全体のリカレント結合のために、スパイクが不安定だが時空間構造に敏感な変化をすると考えられる。この不安定な挙動が、学習メカニズムによって周期的パターンや平均発火率の空間パターンのような安定したダイナミクスや情報表現に変換される過程を、海馬のエピソード記憶を一時的に蓄える処理とみなすことができる。CA3におけるカオス的な不安定性は、現在入力されている情報を柔軟にエンコードするための特性かもしれない。 |