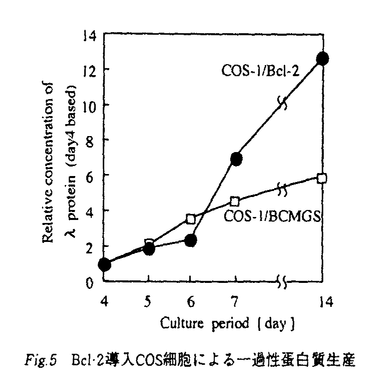
動物培養細胞を用いた蛋白質生産系は、糖鎖付加、プロセッシング等の翻訳後修飾を受けた生理活性の高い蛋白質の生産が可能であり、特に医薬品用蛋白質生産に広く利用されている。しかし、動物細胞を高濃度培地あるいは低血清培地を用いて回分培養すると、培養後期において培地中の微量成分の不足や老廃物の蓄積などが引き金となって、主要な栄養成分が培地中に十分に存在しているにもかかわらず、遺伝子上にプログラムされた死である細胞自殺(アポトーシス)が起こる。その結果、生細胞数が急激に減少し、目的蛋白質の生産もすみやかに停止する。このため、蛋白質の生産期間が短く、また、使用培地量当たりの蛋白質生産量、すなわち蛋白質収率も低くなるという問題点があった。 本論文では、アポトーシス抑制や細胞周期制御に関わる種々の蛋白質の遺伝子を高発現可能なベクターあるいは発現誘導可能なベクターに組み込んで動物培養細胞に導入することにより、アポトーシスを抑制したり、細胞周期を外部から制御可能な細胞株を樹立し、この細胞株を用いた蛋白質生産期間の長期化と生産量の増加を目指した研究の成果を述べており、以下の7章から構成されている。 第1章は序論であり、本研究の背景と目的を述べている。 第2章では、動物細胞培養技術および細胞周期、アポトーシスに関する既往の研究について述べている。 第3章では、遺伝子組み換え蛋白質の工業的生産に広く利用されているチャイニーズハムスター卵母細胞(CHO細胞)を用いた蛋白質生産系において、遺伝子工学的手法によりアポトーシス抑制可能な細胞株の構築を行い、この細胞株のアポトーシス抑制、ならびにマウス抗体 1鎖の生産期間と生産量に関して得られた成果について述べている。すなわち、低血清培地で培養を行った場合、通常のCHO細胞は約2日でアポトーシスが起こり死滅するのに対して、アポトーシス抑制能を持つBcl-2遺伝子を導入したCHO細胞は6日以上生存することが可能となり、その結果、マウス抗体1鎖の生産期間も大幅に長期化することを報告している。一方、同じくアポトーシス抑制能を持つと考えられているp35遺伝子を導入したCHO細胞の場合にはこのようなアポトーシスの抑制効果は観察されず、通常のCHO細胞と同程度の生存期間であったことを報告している。さらに、Bcl-2遺伝子、p35遺伝子を導入したCHO細胞では、通常のCHO細胞と比較して増殖速度は低下するものの、マウス抗体1鎖の比生産速度がそれぞれ4倍、2倍に増加することを明らかにしている。このようにアポトーシス抑制遺伝子の導入により、抗体の生産期間を長期化したり、蛋白質の比生産速度を向上させることにより、蛋白質生産量を2倍程度まで増加させることに成功している。 1鎖の生産期間と生産量に関して得られた成果について述べている。すなわち、低血清培地で培養を行った場合、通常のCHO細胞は約2日でアポトーシスが起こり死滅するのに対して、アポトーシス抑制能を持つBcl-2遺伝子を導入したCHO細胞は6日以上生存することが可能となり、その結果、マウス抗体1鎖の生産期間も大幅に長期化することを報告している。一方、同じくアポトーシス抑制能を持つと考えられているp35遺伝子を導入したCHO細胞の場合にはこのようなアポトーシスの抑制効果は観察されず、通常のCHO細胞と同程度の生存期間であったことを報告している。さらに、Bcl-2遺伝子、p35遺伝子を導入したCHO細胞では、通常のCHO細胞と比較して増殖速度は低下するものの、マウス抗体1鎖の比生産速度がそれぞれ4倍、2倍に増加することを明らかにしている。このようにアポトーシス抑制遺伝子の導入により、抗体の生産期間を長期化したり、蛋白質の比生産速度を向上させることにより、蛋白質生産量を2倍程度まで増加させることに成功している。 第4章では、組み換え蛋白質の一過性発現に広く用いられているCOS細胞にBcl-2遺伝子を導入した細胞株を構築し、この細胞株のアポトーシス抑制、ならびにマウス抗体1鎖の生産期間と生産量に関して得られた成果について述べている。すなわち、低血清培地を用いて培養を行った場合、通常のCOS細胞は全く増殖せず、蛋白質生産も一週間以内で停止するのに対して、アポトーシス抑制能を持つBcl-2遺伝子を導入したCOS細胞の場合には低血清培地のような貧栄養条件下でも増殖することが可能となり、その結果、マウス抗体1鎖の生産が一週間以上継続し、通常のCOS細胞の2倍の蛋白質生産量が得られたことを報告している。一方、アポトーシス抑制効果については、Bcl-2遺伝子を導入した細胞株でも通常のCOS細胞と同様にアポトーシスに特徴的な現象である核内の染色体の断片化が起っており、COS細胞ではBcl-2遺伝子による完全なアポトーシス抑制は困難であると述べている。このように、Bcl-2遺伝子は核内の染色体の断片化は抑制できないものの、蛋白質の生産期間を延長する効果があったこと、さらに、Bcl-2遺伝子産物がミトコンドリアの膜に存在するという他の研究者の報告に基づき、Bcl-2遺伝子産物の過剰発現によりアポトーシスの過程でおこるミトコンドリアの崩壊が抑制され、その結果としてミトコンドリアにおけるATP生産が維持されたことが、蛋白質生産量の増加の原因であると考察している。 第5章では、診断薬、治療薬として注目を浴びているモノクローナル抗体の工業的生産に広く用いられているハイブリドーマ細胞を用いた抗体生産系において、遺伝子工学的な手法により細胞周期制御あるいは増殖速度制御が可能な細胞株の構築を行い、この細胞株の増殖速度制御、ならびに抗体の生産期間と生産量に関して得られた成果について述べている。すなわち、蛋白質生産が最も活発に行われるG1期に細胞周期を停滞させることで蛋白質生産量を増加させることを目標として、G1期において細胞周期の進行を阻害する蛋白質として知られているサイクリンインヒビターp21あるいはp27の遺伝子をハイブリドーマ2E3細胞に導入し、これらの遺伝子を発現誘導させることにより、細胞の増殖速度を低下させ、p21遺伝子を導入した細胞では抗体生産量を通常のハイブリドーマ細胞の生産量の1.5倍に、また、p27遺伝子を導入した細胞では抗体生産量を3倍に増加させることに成功している。p27遺伝子を導入した細胞の場合には、細胞の生存期間は通常の細胞の8日から12日へと長期化され、抗体の比生産速度も通常細胞の1.5倍程度の高い値を培養後期まで維持していたが、一方で細胞の最大到達濃度は通常のハイブリドーマ細胞の約80%とかなり低下した。これは、p27遺伝子誘導発現による細胞周期進行の阻害効果が強すぎたため、G1期停滞のみならず細胞のアポトーシスも誘発したためと考察している。これに対して、p21遺伝子を誘導発現させた場合には、細胞の増殖速度は低下するものの、細胞の最大到達濃度は通常のハイブリドーマ細胞と同等であり、細胞の生存期間は通常の細胞の8日から12日へと長期化された。また、抗体の比生産速度は通常細胞と同程度であったが、培養後期までその生産活性が維持された。これは、p21遺伝子を誘導発現させた場合には、アポトーシスを誘発せずに細胞周期を適度にG1期に停滞させることができたためと考察している。 第6章では、本論文を総括し、ここで構築したアポトーシス抑制、細胞周期制御が可能な細胞株を工業的生産規模での蛋白質生産へ応用する上での展望を述べている。 第7章は結言である。 以上、本論文は、有用蛋白質の工業的な生産に広く利用されている動物培養細胞であるCHO細胞、COS細胞、ハイブリドーマ細胞を対象として、遺伝子工学的手法によりアポトーシス抑制、細胞周期制御可能な細胞株を構築し、アポトーシス抑制、細胞周期制御などの技術により蛋白質生産期間の長期化や生産量の増大が可能であることを実証したものであり、動物培養細胞を用いた蛋白質生産技術の向上に寄与するところが大きい。 よって、本論文は博士(工学)の学位請求論文として合格と認められる。 |