イネの子実に蓄積される炭水化物は、次の二つに由来すると言われている。第一は、出穂前に一旦、稈や葉鞘などの栄養器官に蓄積され、出穂後に穂へ転流されてくるものであり、第二は、出穂後、葉の光合成によって作られたものが直接穂に送られてくるものである。子実収量に対する転流炭水化物の貢献は、品種、あるいは窒素施肥、栽植密度などの栽培条件で異なると言われている。近年、この転流炭水化物のイネ収量への貢献については、イネの収量形成機構に関する研究の重要な課題として研究が進められている。本研究は、イネの収量形成における転流炭水化物と出穂後の光合成に由来する炭水化物の役割を検討したものである。まず第1章において、出穂後における葉や稈などの栄養器官、および穂における炭水化物の変化の実態を調べるとともに、転流炭水化物と出穂後光合成由来炭水化物との穂への転流が、光条件でどのように制御されているかを検討した。第2章においては、印度稲、日本稲、ジャヴァ稲の品種について、出穂後の栄養器官に蓄積された炭水化物のレベルと、その時間的変化を調べた。第3章においては、葉鞘における炭水化物レベルと、デンプンの合成あるいは分解に関わる酵素の活性との関係を調べ、葉鞘における炭水化物レベルがどのような生化学的機構で制御されているかを調べた。最後に第4章においては、供給された炭水化物が穂においてどう使われるか、主に呼吸基質としての使われ方を中心に検討した。得られた結果の概要は次のとおりである。 1.出穂後の葉・稈および穂における炭水化物の消長 出穂後0-22日の、止め葉を含む上位3枚の葉身、葉鞘および穂におけるデンプン、蔗糖の消長を調べた。葉鞘からのデンプン流出は、止め葉を含む上位3枚の葉の葉鞘間で並行的に起こっており、それと同時に、穂においては急激なデンプンの蓄積が観察された。異なる栄養器官でデンプン蓄積レベルを比較してみると、デンプン貯蔵は稈で最も多いことが示された。また、異なる葉位間では、第2、第3葉葉鞘で多く、第1葉(止め葉)葉鞘では低いことが示された。デンプン蓄積が最大になった出穂期でさえも栄養器官におけるデンプン蓄積は最大レベルにまで達しておらず、第2、第3葉葉鞘では最大容量の約78%しか蓄積していなかった。こうしたことから、栄養器官におけるデンプン蓄積は、器官形成に使われた後の余剰の炭水化物が蓄積するものと考えられた。登熟器官においても、栄養器官における炭水化物の蓄積能力は失われていなかった。登熟がほぼ完了した出穂後22日以後には、第1葉葉鞘においてデンプン蓄積が再開し、出穂時の87%のレベルにまで回復した。 一方、蔗糖含量は、第1葉においては葉身と葉鞘の両者でほぼ同じ値を示したが、第2葉、第3葉については葉身の方が葉鞘よりも高い価を示し、葉身が直接の光合成産物の供給源となっていることが示された。また、止め葉葉身の蔗糖含量は、第2、第3葉身よりも高く、止め葉が光合成由来炭水化物の主要なソースとなっていることが示された。一方、穂の蔗糖含量は、いずれの葉の葉身・葉鞘よりも高くなっていた。 次に、蓄積炭水化物と同化炭水化物の穂の生長に対する相対的な関係を調べた。植物体を遮光すると、同化炭水化物の供給が減少するが、それを補償するように、蓄積炭水化物の供給が増加した。また、1日で見ると、昼間は同化炭水化物の供給が主体をなしたが、夜間は蓄積炭水化物の供給が主体をなした。このように、栄養器官に蓄積される蓄積炭水化物の子実収量への貢献は、出穂後の光合成が低下するような条件下で、穂に転流されると考えられた。 2.栄養器官における炭水化物蓄積の品種間差 印度稲、日本稲、ジャヴァ稲各2品種につき、第3葉葉鞘におけるデンプンの出穂前蓄積量を比較した。印度稲の2品種は、高い蓄積レベルを示した。日本稲の2品種は中位の蓄積レベルを示した。ジャヴァ稲の2品種は低い蓄積レベルを示した。このことから、ジャヴァ稲は茎が太く、稈も長いにもかかわらず、デンプンの蓄積量が少ないのは、供給される炭水化物のうち、多くの部分が栄養器官の形成素材として利用されてしまう結果、蓄積される炭水化物は低くなっていると考えられた。しかし、こうしたデンプンの出穂前蓄積量の違いは、子実収量の品種間差には反映されていなかった。そこで、シンク側の要因である収量構成要素を調べてみると、子実収量は、穎果の大きさ、および穎果の数との間に正の相関を示し、子実収量は主にシンク容量によって決定されていることが示された。さらに、出穂後の光合成が低下し、第3葉葉鞘における出穂前蓄積デンプンの穂への転流量が相対的に大きくなると、登熟歩合が低下する現象が認められ、登熟歩合の決定に対しても出穂前蓄積デンプンの役割は比較的小さいと考えられた。 3.栄養器官における糖代謝に関連する酵素活性の時期的変化 登熟期間中の葉鞘におけるデンプン含量、デンプン合成関連酵素活性、デンプン分解関連酵素活性を調べ、相互の関係を検討した。まず、デンプン含量は、第1章でも観察されたように、出穂後7-11日目に増加傾向を示したが、大きな傾向としては出穂後、一貫して減少傾向を示していた。しかしながら、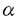 -アミラーゼやスターチホスホリラーゼなどのデンプン分解関連酵素の活性は、これに関連しておらず、出穂後0-22日の期間、ほぼ一定の活性を維持していた。一方、可溶性デンプン合成酵素やADPグルコースピロホスホリラーゼなどのデンプン合成関連酵素活性は、デンプン含量の変化に類似した変化傾向を示していた。これらのことから、葉鞘内に蓄積されるデンプンのレベルは、デンプン合成関連酵素活性の消長によって制御されており、デンプン分解に関わる酵素による制御は弱いと考えられた。ただし、デンプン合成関連酵素のうちQ-酵素は、出穂後長期間にわたって活性が高く維持されていたことから、この酵素による葉鞘内デンプンレベルの制御は弱いと考えられた。 -アミラーゼやスターチホスホリラーゼなどのデンプン分解関連酵素の活性は、これに関連しておらず、出穂後0-22日の期間、ほぼ一定の活性を維持していた。一方、可溶性デンプン合成酵素やADPグルコースピロホスホリラーゼなどのデンプン合成関連酵素活性は、デンプン含量の変化に類似した変化傾向を示していた。これらのことから、葉鞘内に蓄積されるデンプンのレベルは、デンプン合成関連酵素活性の消長によって制御されており、デンプン分解に関わる酵素による制御は弱いと考えられた。ただし、デンプン合成関連酵素のうちQ-酵素は、出穂後長期間にわたって活性が高く維持されていたことから、この酵素による葉鞘内デンプンレベルの制御は弱いと考えられた。 4.穂における炭水化物の利用 穂に供給される炭水化物のうち、昼間に供給されるものは主に光合成から、夜間に供給されるものは主に貯蔵炭水化物に由来していた。穂に供給される炭水化物を穂の呼吸基質に使われる部分と、穂の生長素材として使われる部分とに分けてみた。穂の生長が最も活発な出穂後12日の穂において、全供給炭水化物のうち穂の収量形成に使われたものの割合、すなわち生長効率は約68%であった。逆に言えば、32%が穂で呼吸に使われていた。そこで、呼吸の内容を分析してみた。呼吸は、生長呼吸と維持呼吸とに分類される。測定の結果、生長呼吸は約55%、維持呼吸は約45%を占めていた。維持呼吸はさらに、シアン感受性呼吸とシアン耐性呼吸とに分類される。維持呼吸のうち、シアン感受性呼吸、すなわち電子伝達系を経由する呼吸は76%、(全体の呼吸からすると35%)シアン耐性呼吸、すなわちalternative respirationは約23%(全体の呼吸からすると10%)であった。 |