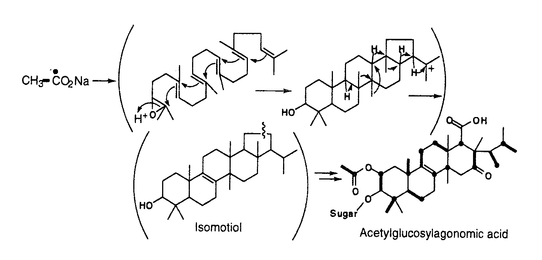
テルペノイドは、C5化合物であるイソペンテニル2リン酸(IPP)の縮合により生成される化合物であり、生体の構成成分として、あるいは生理活性物質などとして多様な役割を果たしている。これらのテルペノイドの生合成には未解決な問題が多く存在し、その解明は学術的に重要であるとともに、様々な分野における物質生産の基礎ともなるものである。 本論文はこのような背景に基づき、特に多様性に富み、産業的応用など広範囲の可能性を有する微生物の生産するテルペノイドの生合成研究を行って、様々な新事実を明らかにしたものであり、3章よりなる。 第1章は、アセチルグルコシルアゴノミシ酸(WF11605)の生合成研究について述べている。13C標識された酢酸の添加により調製したサンプルのNMR解析によって、本物質はスクアレンを経て生合成されることを明らかにした。また、13CH313CO2Naで標識されたサンプルのTANGO-HMBCスペクトルから、メチル基の転位反応を直接証明することにはじめて成功した。 第2章は、放線菌の生産するテルペノイドの生合成に関するものである。まずナフテルピンの生産菌であるStreptomyces aeriouviferにおけるテルペノイドの生合成研究について述べている。13C標識された酢酸とグルコースを添加培養し、一次代謝産物であるメナキノンと二次代謝産物であるナフテルピンの標識パターンの解析を行った。その結果S.aeriouviferにはメバロン酸経路および非メバロン酸経路の両方の経路が存在し、生育初期に生産されるメナキノンは前者で、後期に生産されるナフテルピンは後者で主に生合成されるという、興味深い事実を明らかにした。 本章の後半では、放線菌におけるメバロン酸経路について述べている。S.aeriouviferのHMG-CoAレダクターゼ遺伝子をプローブとするサザンハイブリダイゼーションによる解析法を開発し、この方法がメバロン酸経路の存在の証明に有用であることを示した。また、本方法の応用により、メバロン酸経路を有する放線菌の分布が稀であることが明らかとなった。 第3章では、非メバロン酸経路の解析について説明している。まず最初に2-C-メチル-D-エリスリトール4-リン酸(MEP)の生合成機構に関して記述している。ランダムに変異を導入した大腸菌から、合成した2-C-メチルエリスリトール(ME)添加時のみ生育できるMEP合成遺伝子欠損株をスクリーニングした。次いで、大腸菌のゲノムライブラリーから、この変異を相補する遺伝子のクローニングを行った。さらに、この遺伝子産物の機能を明らかにするため、この遺伝子の大量発現を行って得られたタンパク質をMn2+とNADPHの存在下、1-デオキシキシルロース5-リン酸(DXP)と反応させ、生成物を精製して解析したところ、MEPであることが判明した。この結果から、このタンパクはDXP骨格の分子内転位と同時に還元反応をも触媒し、一段階でDXPからMEPを合成する酵素であることが明らかとなり、DXPレダクトイソメラーゼ(DXR)と命名した。 ついでMEP以降の非メバロン酸経路について説明している。DXR遺伝子破壊株に合成した[1,1-2H2]2-C-メチル-D-エリスリトールを添加培養して得られたメナキノンを解析したところ、MEPの1位の2つのプロトンは完全に保持されてIPPに取り込まれることが判明し、反応機構に付いての重要な情報が得られた。 以上本論文は、微生物の生産するテルペノイドの生合成研究を行い、様々な新事実を明らかにしたものであって、学術上、応用上寄与するところが少なくない。よって、審査委員一同は、本論文が博士(農学)の学位論文として価値あるものと認めた。 |