現在、世界の人口増加とその人間活動に基づく地球環境の破壊が深刻な食糧不足をもたらすであろうことが多くの研究者により指摘されている。食糧不足への対策としては農耕地の拡大、作物生産性の向上をはかることが不可欠であるが、地球上の陸地の3分の1は農耕に不適格な乾燥地および半乾燥地であり、また世界の主要穀倉地の10%以上が過度の潅漑に伴う塩類集積により著しい収量減産を招いている。こうした乾燥や塩類集積などの環境ストレスを解決する有力な方法として、一つは耐乾燥性や耐塩性の強い品種を選抜していくこと、もう一つは近年発展の著しいバイオテクノロジーの手法を用いて作物に耐乾燥性や耐塩性遺伝子を導入することにより優れた耐性品種を作出することがある。しかし生物の環境ストレス耐性はかなり多数の機能から構成されており、それぞれの種における各機能の組合せも単純ではなく、解明しなければならない問題を極めて多く含んでいる。 上記を踏まえ、本研究では、様々な環境ストレスのうち塩ストレスに焦点を絞り、微生物を用いて耐塩性と浸透圧調節機構との関係を解明することを目的として、以下の観点から研究を行った。 1.塩ストレスに対する細胞内の物質的応答の探索 耐塩性を持つ生物では、細胞が浸透圧ストレスを受けた時に、細胞内浸透圧の上昇を防ぐ役割を果たす低分子有機化合物を細胞内に多量に蓄積することが知られている。このような物質を適合溶質と呼ぶ。また無機イオンの中にも適合溶質と同様の機能を持つものがあると考えられている。そこで本章では、脱窒光合成細菌Rhodobacter sphaeroides f.sp.denitrificans IL 106を異なる塩濃度培地で培養し、菌体内の無機イオンならびに可溶性有機成分の変化を追跡した。本菌は淡水性細菌でありながら海水と同程度の塩濃度の培地でも生育し、比較的強い耐塩性を示すことがすでに明らかにされている。 菌体内の無機イオンを炎光法及びイオン交換クロマトグラフィーで測定したところ、培地の塩濃度の上昇に伴って細胞内のカリウムイオン濃度が顕著に増加し、一方ナトリウムイオンや塩化物イオンの増加は抑制されていた。このことから、カリウムイオンは細胞内の浸透圧バランスをとる役割を果たしているのではないかと推察された。 次に可溶性有機成分の変化をイオン交換クロマトグラフィーおよびNMRで追跡した。培地中のNaCl濃度が4%の場合、試料の1H-および13C-NMRスペクトルからは六炭糖が一種類存在することが示され、この糖は還元末端を持たないことからアノメリック炭素同士が結合した二糖であると考えられた。詳細なスペクトル解析の結果、この条件を満たす糖としては唯一トレハロースが考えられ、標品と比較した結果完全に一致した。これにより、低分子有機適合溶質として、きわめて多量のトレハロースが細胞内に誘導合成されること、さらにその誘導が培地塩濃度依存的であることを明らかにした。 細胞内における主要な浸透圧調節物質の存在量を定量し、その比を求めた結果、0%NaCl培地の場合に比べ4%NaCl培地では、カリウムイオンとトレハロースとの合計モル数が約6倍強に増加した。このことから、無機イオンとしてはカリウムイオンが、有機適合溶質としてはトレハロースが主要な浸透圧調節物質であることを証明した。他に、遊離アミノ酸も培地塩濃度の上昇とともに若干増加するが、存在量がかなり低く、適合溶質としては寄与していないと考えられた。 2.塩ストレス誘導タンパクの探索およびその生化学的性質の解析 耐塩性微生物は塩ストレス下で、適合溶質を蓄積する以外にも、さまざまな代謝系に関与するタンパク質を誘導合成することにより細胞内浸透圧を調節しているものと考えられている。そこで本章では、R.sphaeroidesにおいて塩ストレス下誘導合成されるタンパク質をSDS-PAGEにより調べた。非ストレス条件下では微量しか認められない分子量39kDaのタンパク質が、塩ストレス下で細胞膜に集積するのが認められた。またペリプラズムのバンドのパターンにも若干の変化が見られたが、細胞質にはほとんど変化が認められなかった。膜画分をさらに分画して同様の実験をしたところ、39kDaタンパク質は細胞質膜ではなく外膜に蓄積することが判明した。 次に、塩ストレスによって存在量が大きく変化するタンパク質を二次元電気泳動により確認した。0%NaCl培地の場合と4%NaCl培地の場合とを比べた結果、前述の39kDaタンパク質と思われるスポットSsp1(MW39kDa,pI5.0)が膜画分において確認された。さらに変化するスポットを2つ(Ssp0,Ssp2)発見した。Ssp0(MW52kDa,pI5.1)は塩ストレス下で減少し、Ssp2(MW50kDa,pI5.2)は増加した。ペリプラズム画分においては、7スポットに変化が見られた。そのうちの1スポットは4%NaCl培地の場合に減少したが、それ以外のスポットはすべて増加した。 二次元電気泳動により存在量の変化が確認された3種類の膜タンパク質(Ssp0,Ssp1,Ssp2)について、それぞれアミノ酸配列解析を行った。その結果、Ssp0のN末端アミノ酸配列はflagellar filament proteinと一致し、Ssp2のN末端アミノ酸配列はATP synthase beta chainと一致した。39kDaのSsp1はアミノ酸配列が読めなかったことから、N末端がブロックされていると推測した。そこでSsp1をトリプシンあるいはStaphylococcus aureus V8プロテアーゼで断片化することにより4つのペブチド断片を得た。それらのアミノ酸配列をデータベースで検索したところ、有意な相同性を持つタンパク質は認められなかったため、Ssp1は新規タンパク質であることが強く示唆された。 塩ストレスにより最も多く誘導された39kDa外膜タンパク質Ssp1に焦点を絞り、生化学的性質を解析した。このタンパク質は高塩濃度条件や高pH条件では可溶化されず、界面活性剤Triton X-100により可溶化されたことから、膜貫通型タンパク質であると推定された。熱ショック、塩ストレス、乾燥ストレス及び低温ストレスなどの水ストレスで共通に誘導されるタンパク質は多いが、このタンパク質は熱ショックでは誘導されなかったので、塩ストレスに特有の誘導タンパク質である可能性が示唆された。さらに、対角線電気泳動法によりS-S結合が存在していないことが推定された。 3.誘導タンパク質Ssp1の遺伝子クローニングとアミノ酸配列解析 新規タンパク質であることが示唆された39kDa外膜タンパク質の遺伝子クローニングを行った。まず、アミノ酸配列が最も長く読めた18アミノ酸残基に対応するDNA断片(53bp)をR.sphaeroidesゲノムからPCR法により増幅し、プラスミド上にクローン化した。次に、この53bpのDNA断片をプローブとしたサザンハイブリダイゼーションから、目的遺伝子の少なくとも一部は2.6kbのPstI DNA断片上に存在することが明らかになった。そこでこの大きさのPstI DNA断片をアガロースゲルから抽出精製し、クローニングベクター上に導入した後、これを用いて、大腸菌を形質転換し、約1000個の形質転換体を得た。これらの形質転換体についてPCR法によるスクリーニングを行ったところ、目的の挿入断片をもつ陽性クローンがひとつ得られた。その一次配列を決定したところ、359アミノ酸残基からなるタンパク質をコードしうるオープンリーディングフレームが見いだされた。この予想されるアミノ酸配列は、前章においてプロテアーゼ処理断片から決定した4つの部分アミノ酸配列の全てを含んでいた。配列から予想される分子量は39.5kDa、等電点は4.66であり、これは39kDa膜タンパク質のものとほぼ一致した。データベース検索の結果、有意な相同性を持つタンパク質は認められなかったため、このタンパク質は塩ストレスによって誘導される新規タンパク質と確認された。現在、その機能の解明を行っている。 まとめ 以上を要約すると次の通りである。 1)光合成細菌Rhodobacter sphaeroidesにおいて、塩ストレス下で細胞内に無機イオンとしてカリウムイオンが、また有機適合溶質としてトレハロースが蓄積され、さらに、その蓄積が培地塩濃度依存的であることを示した。 2)本菌において塩ストレス下で数種類のタンパク質の誘導あるいは欠失が認められた。塩ストレスにより最も多く誘導された39kDa外膜タンパク質の細胞内局在性を明らかにし、いくつかの生化学的性質を調べた。 3)誘導タンパク質Ssp1は、遺伝子クローニングの結果、新規タンパク質であることが判明した。 | 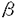 chainであった。最も誘導量の多いSsp1はN末端配列がブロックされており、配列が読めなかった。そこでSsp1をトリプシンおよびStaphylococcus aureus V8プロテアーゼで断片化した。得られた4つの断片のアミノ酸配列をデータベースで検索したところ、有意な相同性を持つタンパク質は認められなかったため、Ssp1は新規タンパク質であることが強く示唆された。
chainであった。最も誘導量の多いSsp1はN末端配列がブロックされており、配列が読めなかった。そこでSsp1をトリプシンおよびStaphylococcus aureus V8プロテアーゼで断片化した。得られた4つの断片のアミノ酸配列をデータベースで検索したところ、有意な相同性を持つタンパク質は認められなかったため、Ssp1は新規タンパク質であることが強く示唆された。