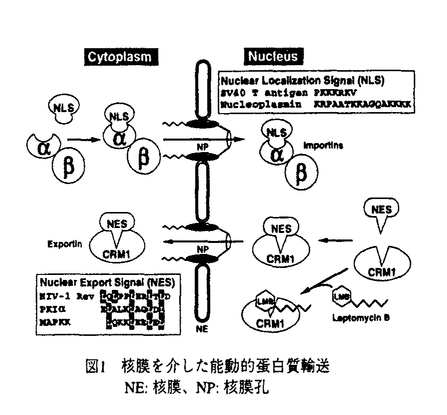
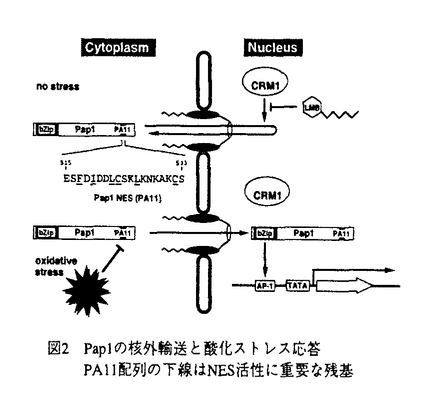
レプトマイシンB(LMB)はnMオーダーの低濃度で分裂酵母、ラット正常線維芽細胞3Y1に対しG1、G2期特異的な細胞周期の停止を、SV40形質転換細胞に対してはアポトーシス様細胞死を誘導するとともに実験動物がんに対する抗腫瘍活性を示す放線菌の代謝産物である。本物質の標的分子は以前の分裂酵母の遺伝学的解析によりcrm1+遺伝子産物であることが示唆されていたが、その機能は不明であった。レプトマイシンの標的分子CRM1蛋白質の機能に関する研究と題した本研究は、上に述べたようなユニークな活性を示すLMBの標的と考えられるCRM1蛋白質の機能を解明することを目的として、そのヒト・ホモログのクローン化、CRM1とLMBの相互作用の解明等を通じてCRM1が真核生物に共通の蛋白質核外移行の受容体であることを証明し、さらにそれがストレス応答に関与することを明らかにし、シグナル伝達の新たな分子機構を明らかにしたものであり、6章から構成される。 研究の背景と意義を述べた第1章に続き、第2章ではヒトCRM1のcDNAを新たに単離し、発現、細胞内局在に関する解析を行った。取得したCRM1の機能的なヒトホモログ(hCRM1)は全長にわたり酵母CRM1と極めて高い相同性を示した。hCRM1の細胞内局在を解析し、主として核膜に局在することが明らかとなり核膜機能と関連する蛋白質であることが予想された。また、CRM1はG1後期に転写が始まり、G2/M期にピークになるという細胞周期依存的な発現調節を受けることも明らかになった。 第3章では、LMBの標的分子が真にCRM1であるかを明らかにするために、生化学的に細胞からLMB結合蛋白質を同定した。細胞抽出液からLMBのビオチン化誘導体と結合する蛋白質を検出し、この蛋白質がhCRM1であることを示し、LMBの標的分子がCRM1であることを証明した。さらに、分裂酵母のLMB耐性化株の遺伝学的解析をもとにして、LMBがCRM1のCys-529を介し共有結合することが明らかになった。 第4章では、CRM1蛋白質の機能を分裂酵母と動物細胞で解析し、CRM1がnuclear export signal(NES)依存的蛋白質核外移行の受容体であることを証明した。まず分裂酵母で核-細胞質間蛋白輸送を解析する系を構築しLMBの効果とCRM1の関与について解析を行った。その結果、LMB処理とCRM1の低温感受性変異株でNES依存的な蛋白質核外移行が特異的に阻害され、このことから、分裂酵母においてCRM1の機能は蛋白質の核外移行に必須であることが示された。動物細胞におけるマイクロ・インジェクションを用いた実験でも同様な結論が導かれた。さらに、NESとCRM1の結合を生化学的に示し、その結合はLMBによって阻害されることが明らかとなった。これらの結果から、CRM1は真核細胞全般に保存されたNESの受容体であり、LMBはCRM1とNESの結合を阻害することによって蛋白質核外移行を阻害することが非常に明確に示された。 第5章では、分裂酵母AP-1様転写因子Pap1の酸化ストレス応答性核外移行シグナルの解析を通じ、細胞のストレス応答の新たな分子機構の発見について述べられている。分裂酵母の転写因子Pap1の活性はCRM1によって負に制御されていることが予想されていたことから、Pap1の細胞内局在を調べたところ、細胞質だけに局在しLMBの添加とCRM1の失活で核移行したことから、Pap1はCRM1を介し核外移行されることが明らかとなった。次にPap1のNESを同定したところ、C末端領域にPA11領域を見出した。さらに酸化ストレス条件に応答しPap1が核移行することを明らかにした。その応答は一般的なNESと異なりPap1 NESに独特な機能であった。さらに、Pap1のNESは高等真核細胞でも機能することが明らかになった。以上のPap1の解析から、真核細胞全般に保存された新たなストレス応答の分子機構を発見した。 第6章の総括では、本研究のまとめと成果の意義、今後の研究への応用について論じている。 以上、本論文では、LMBの標的分子が真にCRM1蛋白質であることを明らかにした上で、そのCRM1蛋白質がこれまで未同定だったNES依存的蛋白質核外移行の受容体、すなわち「exportin」であることを解明した。さらに、転写因子Pap1の制御機構の解析から、これまでに全く知られていなかった蛋白質の核外移行を介した酸化ストレス応答の分子機構の存在を発見したもので、学術上貢献するところが少なくない。よって、審査委員一同は本論文が博士(農学)の学位論文として価値のあるものと認めた。 |  g/ml)を取得した。同株のLMB高度耐性変異は遺伝解析によりcrm1変異と連鎖し、crm1遺伝子自身の変異であると予想された。そこでFN41株のcrm1遺伝子を単離し解析したところ、Cys-529のSerへの唯一の変異がLMB高度耐性化に十分であることが明らかになった。興味深いことに、分裂酵母同様LMB感受性のヒトCRM1では対応する残基はCysであるが、LMB非感受性である出芽酵母ではThrである。このことからもこのCys残基はLMBの共有結合部位であることが予想された。実際、マス・スペクトル解析の結果、該当するCys残基を含むhCRM1のペプチド(18アミノ酸残基)に対しLMBが不飽和ラクトン部分を介してマイケル付加されることが示されたことから、Cys-529がLMBと共有結合するアミノ酸残基であることが示唆された。
g/ml)を取得した。同株のLMB高度耐性変異は遺伝解析によりcrm1変異と連鎖し、crm1遺伝子自身の変異であると予想された。そこでFN41株のcrm1遺伝子を単離し解析したところ、Cys-529のSerへの唯一の変異がLMB高度耐性化に十分であることが明らかになった。興味深いことに、分裂酵母同様LMB感受性のヒトCRM1では対応する残基はCysであるが、LMB非感受性である出芽酵母ではThrである。このことからもこのCys残基はLMBの共有結合部位であることが予想された。実際、マス・スペクトル解析の結果、該当するCys残基を含むhCRM1のペプチド(18アミノ酸残基)に対しLMBが不飽和ラクトン部分を介してマイケル付加されることが示されたことから、Cys-529がLMBと共有結合するアミノ酸残基であることが示唆された。