発生初期における神経回路綱の形成過程では、まず過剰の神経細胞が産生され、その後、標的組織と機能的シナプス結合を形成できなかった神経細胞は、選択的細胞死によって除去される。この選択的細胞死は、中枢神経系(脳、脊髄)および末梢神経系の多くの神経細胞において認められ、標的組織由来の栄養因子を獲得できなかったために起こると考えられている。本研究は、脊髄運動神経に対して生存維持活性を持つ物質を、その標的器官である骨格筋から精製し、構造解析を行なったもので、8章からなっている。 研究の背景と意義を述べた第1章に続き、第2章ではニワトリ6日胚の脊髄神経細胞初代培養系を用いた生物検定法を述べている。この生物検定法では、骨格筋抽出液など生存維持活性を示す物質を培地中に加えると、脊髄神経細胞は2日間生存して神経突起を伸展させたが、何も加えないとすべての細胞が2日以内に死滅した。 第3章では胚期後期18-19日胚の骨格筋からの活性物質精製法について述べている。アセトンによる脱脂、2%食塩水による抽出、アセトンおよび硫安による分画沈澱、さらに2段階のカラムクロマトグラフィーを行い、回収率約20%で部分精製物を得ることに成功した。また、96℃、5分間の熱処理によって完全に失活すること、還元剤や界面活性剤などに不安定であること、ゲルろ過の溶出位置から分子量が17000-47000と推定されたことなど、活性物質の基本的性質を明らかにした。引き続き、部分精製物を陰イオン交換HPLCおよび疎水性クロマトPOROS50PEによって精製し、さらに逆相HPLCでブロードではあるが、単一なピークとして活性物質を回収した。 第4章では最終精製物が258nmに極大吸収を示し、A260/A280の比率は1.9であること、さら1H-NMR測定の結果、高磁場側から順にリボースのH2’,H3’,H4’,H5’,H5"に帰属される重なり合ったシグナル、およびリボースのH1’とピリミジン環のH5の重なったシグナル、およびプリン環のH8とピリミジン環のH6の重なったシグナルがそれぞれ観測されたことから、リボヌクレオチドを構成単位とするポリマー、すなわちRNAである可能性が高いことを明らかにした。 次に、RNAが神経生存維持活性を担っている物質であることを確かめるため、核酸抽出の常法であるフェノール・クロロホルムで抽出、また、DNase、ProteinaseKおよびRNaseAの3種類の酵素処理による失活実験を行った。その結果、フェノール・クロロホルムで抽出を行っても活性が完全に維持されること、DNase処理では活性は維持され、ProteinaseK処理ではやや失活するものの、RNaseA処理ではほぼ完全に失活することを明らかにし、骨格筋抽出物中に含まれる神経生存活性を示す物質はRNAであると結論した。第5章では、活性物質がRNAであることから、フェノール・クロロホルム抽出など核酸精製に有効な方法を加え、効率の良い精製法を確立した。 第6章では活性物質であるRNAの配列を明らかにするために化学修飾分解法を用いて、塩基配列の決定を試みた。まず、最終精製物の純度を確かめるために、[5’-32P]-pCpを3’末端に導入した後、ポリアクリルアミド電気泳動を行なった。その結果、160bpの2本鎖DNAマーカー近辺にバンドが11本認められ、最終精製物が単一なRNAでないことが分かった。そこで、各々のバンドを切り出し、再度、電気泳動を行うことで純度を高め、このうちの4本のバンドについて塩基配列を決定した。解析の結果、3’末端から約20塩基については配列を明らかにすることができたが、他の部分については確実な配列を明らかにすることはできなかった。その原因はRNA中に修飾塩基が含まれているために、その塩基部位で化学反応および酵素切断が起きなかったことが一因であると考えた。さらに、明らかになった配列から3本のバンドは、同じRNA分子から3’側、または5’側が部分的に欠失したRNAであると考えられた。また、第2章で精製した活性物質のいくつかのバンドについて塩基配列を決定したところ、同じ部分配列が得られた。以上の結果から、胚期骨格筋中のある特定の配列をもったRNAが脊髄神経細胞の生存維持活性を有しているものと結論された。 第7章は総合討論で本研究のまとめと今後の展望を、また、第8章は実験方法を述べている。 以上本論文は骨格筋由来の運動神経生存因子を精製し、この因子がある特定の配列をもったRNAであることを示したもので、学術上貢献するところが少なくない。よって審査委員一同は、本論文が博士(農学)の学位論文として価値あるものと認めた。 | 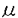 gであり、神経生存維持活性に必要な最小濃度は分子量を47,000とすると0.33nMであった。ところで、最終精製物が単一なRNAであるかどうかを確かめるために、溶出溶液のpHを6.5に変え、同じCapcell-Pak C18による分析を行った。その結果、活性はブロードなピークの肩にのみ認められ、最終精製物中には複数の物質が含まれていることが示された。しかしながら、UV吸収から含まれる物質はいずれもRNAであると考えられた。また、この段階で活性が極端に減少したため、これ以上の精製を行うのは困難であった。一方、あらかじめDNase処理をした試料をポリアクリルアミドゲルを用いて電気泳動したところ、2本鎖DNAマーカーで70bpから160bpの間に約20本のバンドが認められた。さらにこれらを切り出して活性を調べたが、失活が著しく、どのバンドが活性本体であるか不明であった。
gであり、神経生存維持活性に必要な最小濃度は分子量を47,000とすると0.33nMであった。ところで、最終精製物が単一なRNAであるかどうかを確かめるために、溶出溶液のpHを6.5に変え、同じCapcell-Pak C18による分析を行った。その結果、活性はブロードなピークの肩にのみ認められ、最終精製物中には複数の物質が含まれていることが示された。しかしながら、UV吸収から含まれる物質はいずれもRNAであると考えられた。また、この段階で活性が極端に減少したため、これ以上の精製を行うのは困難であった。一方、あらかじめDNase処理をした試料をポリアクリルアミドゲルを用いて電気泳動したところ、2本鎖DNAマーカーで70bpから160bpの間に約20本のバンドが認められた。さらにこれらを切り出して活性を調べたが、失活が著しく、どのバンドが活性本体であるか不明であった。 -32P]-dCTP存在下で65℃にて逆転写を行なったが、逆転写産物は認められなかった。
-32P]-dCTP存在下で65℃にて逆転写を行なったが、逆転写産物は認められなかった。