カルパインは高等動物の細胞質に存在し、Ca2+によって活性化されるシステインプロテアーゼである。細胞内のCa2+によるシグナル伝達に関わり、細胞内で重要な役割をしていると考えられている。近年カルパインはプロテアーゼドメインの相同性から,スーパーファミリーを構成することが明らかとなってきたが、それらの多くはmRNAのレベルで発見されてきたものであり、タンパク質レベルでの解析は遅れている。また、他のプロテアーゼの立体構造が明らかとなるなか、カルパインは立体構造レベルでの研究が依然遅れている。本論文ではバキュロウイルスの大量発現系を用いて、カルパインの構造と機能の相関について解析を行ったものである。 論文の構成は「序論」、「材料」、「方法」、「結果と考察」、「総合討論」、「参考文献」よりなっている。「序論」ではカルパインの歴史、カルパインの活性化機構など、本論文の研究に至った背景および研究目的について述べている。 「結果と考察」の章は「カルパインのバキュロウイルス大量発現系の確立」、「カルパインの結晶化」、「mCL/2’の発現、精製・性質」、「nCL-2’の発現、精製・性質」の4項に分かれている。第1項の「カルパインのバキュロウイルス大量発現系の確立」では組織普遍的に発現している主要なカルパインの1つであるm-カルパインをバキュロウイルス系で発現させた。大腸菌の系ではm-カルパインは不活性な封入体タンパク質として発現するが、変性可溶化後、活性のある状態に再生するのは非常に困難であったので、バキュロウイルスの系を使用した。組み換えm-カルパインは酵素学的に天然のものと同等であり、バキュロウイルス系がカルパイン発現系として非常に有用であることが明らかとなった。次にこの系を用いて、活性中心のシステインをセリンに変換したm-カルパイン(m-C105S-カルパイン)の性質を調べた。プロテアーゼ活性は全く検出されなかった。また、Ca2+存在下で自己消化活性はないものの、構造変化を起こして沈殿することが明らかとなった。この変異体は自己消化をしないのでCa2+存在下での構造などを研究する材料として非常に有用であると考えられる。 第2項では、現在カルパイン研究の主要なテーマであり、非常に競争の激しい分野であるカルパインの結晶化について述べている。カルパインの構造は、カルパインの活性化機構の解明に重要であるにも関わらず、その全体構造は依然明らかとなっていない。本研究では、2.5Åの解像度を示すm-カルパインの結晶をうることに成功した。これは、結晶化に270mgの大量のカルパインを投入した結果であり、「カルパインのバキュロウイルス大量発現系の確立」無くしては、なし得なかったものである。 第3項では、第1項で確立した方法を用いて普遍的なm-カルパイン大サブユニット(m80)の様々な断片のウイルスを作製し、発現を行った。そのうちの一つのmCL/2’は、組織特異的カルパインnCL-2’に相当する断片で、アミノ酸の相同性は71%と高く、まだタンパクレベルで同定されていないnCL-2’のモデルタンパクであることが予想された。精製標品の性質を調べた結果、Ca2+結合ドメインを欠くにも関わらずCa2+依存的な自己消化と基質分解活性が検出された。 第4項では、まだタンパク質レベルで同定されていない胃に特異的に発現しているnCL-2’について、第1項で確立した方法を用いて解析を行った。nCL-2’は1次構造の解析から5つのEF-ハンドからなるCa2+結合ドメインを持たないが、プロテアーゼドメインのすぐ後ろに1つのEF-ハンドの存在が知られていた。本研究でnCL-2’についてCa2+依存的な自己消化活性を確認したことはプロテアーゼドメインの直後のEF-ハンド構造がCa2+を結合することを示唆している。mCL-2’とnCL-2’が活性を保持する形で発現できたので、胃に特異的なnCL-2’の基質の検索やカルパインの生理機能の解明の有効な手段が得られたことになる。 総合討論では以上をふまえ、カルパイン大量発現系の開発と利用、ならびに胃特異的カルパインnCL-2’について総合的な考察を行っている。 以上、本論文は、カルパインの大量発現系を確立し、大量に調製したカルパインを使ってカルパインを結晶化し、結晶解析の基盤を確立すると共に、構造と機能の相関を解析したものであり、学術上貢献するところが少なくない。よって、審査委員一同は本論文が博士(農学)の学位論文として価値のあるものと認めた。 | 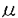 -カルパインの発現が報告されているが、活性化に必要なCa2+感受性が異なるもう一つの主要な組織普遍的カルパインであるm-カルパインの発現は報告されていない。そこでm-カルパインに着目することにし、バキュロウイルスを用いたカルパインの大量発現系を確立した。次にバキュロウイルスのカルパイン発現系を用い、変異型カルパイン、組織特異的カルパイン等を発現・精製し、精製標品の様々な性質を解析した。
-カルパインの発現が報告されているが、活性化に必要なCa2+感受性が異なるもう一つの主要な組織普遍的カルパインであるm-カルパインの発現は報告されていない。そこでm-カルパインに着目することにし、バキュロウイルスを用いたカルパインの大量発現系を確立した。次にバキュロウイルスのカルパイン発現系を用い、変異型カルパイン、組織特異的カルパイン等を発現・精製し、精製標品の様々な性質を解析した。
