 -hexachlorocyclohexane(-HCH、-BHC、lindane)は塩素系の有機農薬であり、広い殺虫スペクトルや非常に安価なことなどから世界中で広く使用されてきたが、人体への有害性、環境中での残留性などが明らかになったため、現在では先進国においてはその製造、使用が禁止されている。しかし、発展途上国などでは経済的理由などからいまだに製造、使用されており、使用された-HCHは揮発した後、気流に乗って世界中に拡散していることが報告されている。Sphingomonas(Pseudomonas)paucimobilis SS86は本学土壌学研究室において単離された-HCHを好気的に分解する報告例の少ない細菌であり、-HCH分解代謝経路については、我々の研究室(細胞遺伝学研究室)においてそのナリジキシン酸耐性株UT26を用いて研究が行われている。これまでの研究で-HCHは2種類の脱クロル酵素LinA、LinBによって2,5-dichloro-2,5-cyclohexadiene-1,4-diol(2,5-DDOL)にまで変換され、さらに2,5-DDOLは脱水素酵素LinCによって2,5-dichlorohydroquinone(2,5-DCHQ)に変換されることが明らかにされている。本研究ではUT26における-HCH分解代謝経路のうち2,5-DCHQ以降の分解に関与する酵素遺伝子の単離と解析をおこなった。 -hexachlorocyclohexane(-HCH、-BHC、lindane)は塩素系の有機農薬であり、広い殺虫スペクトルや非常に安価なことなどから世界中で広く使用されてきたが、人体への有害性、環境中での残留性などが明らかになったため、現在では先進国においてはその製造、使用が禁止されている。しかし、発展途上国などでは経済的理由などからいまだに製造、使用されており、使用された-HCHは揮発した後、気流に乗って世界中に拡散していることが報告されている。Sphingomonas(Pseudomonas)paucimobilis SS86は本学土壌学研究室において単離された-HCHを好気的に分解する報告例の少ない細菌であり、-HCH分解代謝経路については、我々の研究室(細胞遺伝学研究室)においてそのナリジキシン酸耐性株UT26を用いて研究が行われている。これまでの研究で-HCHは2種類の脱クロル酵素LinA、LinBによって2,5-dichloro-2,5-cyclohexadiene-1,4-diol(2,5-DDOL)にまで変換され、さらに2,5-DDOLは脱水素酵素LinCによって2,5-dichlorohydroquinone(2,5-DCHQ)に変換されることが明らかにされている。本研究ではUT26における-HCH分解代謝経路のうち2,5-DCHQ以降の分解に関与する酵素遺伝子の単離と解析をおこなった。 1.linD遺伝子の単離と解析 UT26の遺伝子ライブラリーを広宿主域コスミドベクターpKS13を用いてE.coli HB101、P.putida PpY101中に構築した。それぞれ1,000クローンずつについて2,5-DCHQ分解活性(LinD活性)を調べたところ、P.putidaのライブラリーから6個の陽性クローンが取得された。これらの陽性クローンが保持するコスミドの活性領域を縮小化し、塩基配列を決定したところ、1,038bpのORFがLinD活性に関与していることが明らかとなり、このORFをlinDと命名した。LinDを高発現させた大腸菌に基質である2,5-DCHQを与え、基質の変化を薄層クロマトグラフィー、GC-MSを用いて解析した結果、2,5-DCHQはLinDによってchlorohydroquinone(CHQ)を経てhydroquinone(HQ)にまで変換されることが明らかとなった。相同性検索の結果、LinDはglutathione S-transferaseと相同性を持つことが明らかとなり、LinDを大量発現させた大腸菌の粗酵素抽出液のLinD活性はglutathione存在下で上昇することが判明した。また、linD遺伝子の発現は2,5-DCHQ存在下で誘導されることがlinDをプローブとして用いたUT26全RNAに対するノーザン解析によって明らかとなった。 2.UT26のTn5由来変異株の解析 以前の研究でlinD取得のために、UT26のTn5由来変異株が3株取得されていた。これらの3株、すなわちUT102、UT103、UT116はそれぞれLinD活性について異なる表現型を示すが、挿入されたTn5の近傍にはLinD活性を保持する酵素遺伝子は存在しないことが明らかになっている。つまり、これらの変異株の変異の原因はTn5の挿入ではないことが判明している。これらの変異の原因を解析するため、linD及びその周辺領域をプローブとして用いたサザン解析をUT26及び各変異株の全DNAに対しておこなった。その結果、UT102ではlinDの上流、UT103ではlinD遺伝子中にそれぞれ何らかの断片が挿入されていること、UT116はlinD及びその周辺領域が欠失していることが判明した。UT102、UT103の挿入配列の塩基配列を決定したところ、UT102、UT103に挿入されているのは同一の配列であることが明らかになった。この配列は、両端にUT26のゲノム由来の4bpとそれに続く16bpのinverted repeatを持つ1,654bpの配列で、その中にはIS1380グループのtransposaseと相同性を持つ1,368bpのORFが存在している。サザン解析の結果、この配列はUT26が元々保持している挿入配列であることが明らかとなった。変異株の変異の原因はこの配列がTn5導入時の刺激でlinD及びその周辺領域に転移したためと考えられる。 3.linE遺伝子の単離と解析 LinDは2,5-DCHQをCHQに変換し、さらにHQにまで変換するが、CHQからHQの反応は2,5-DCHQからCHQへの反応に比べて非常に遅い。そのため、UT26中にlinD以外のCHQ分解関与遺伝子が存在する可能性が考えられた。これを調べるため、linDに挿入配列が挿入されている変異株UT103のCHQ分解活性を測定した。その結果、UT103はUT26と同程度のCHQ分解活性を保持していることが明らかとなり、linD以外のCHQ分解関与遺伝子(linE)の存在が示唆された。linEがlinDの近傍に存在している可能性を考え、linD単離時に得たLinD活性陽性クローンのCHQ分解活性を測定したところ、6株中2株で強いCHQ分解活性が観察された。活性領域の縮小化の後に塩基配列の決定を行ったところ、linDの上流約3kbに963bpのORFが同定され、このORFをlinEと命名した。相同性検索の結果、LinEは芳香環のメタ開裂酵素と相同性があることが示唆されたので、LinEと種々のメタ開裂酵素とのアライメントを作製したところ、全体的な相同性は低いものの、活性中心を含めた活性に重要なアミノ酸がほぼ保存されていた。LinEを大量発現させた大腸菌にCHQ、HQを与えて振盪培養し、生成産物を抽出、TMS化の後にGC-MSを用いて同定したところ、CHQは -ketoadipateに、HQは-hydroxymuconic semialdehydeにそれぞれ変換されることが判明した。これにより、LinEは既知のカテコール型の基質(1,2位に水酸基がついている)を開裂するメタ開裂酵素とは異なり、ハイドロキノン型の基質(1,4位に水酸基がついている)を開裂する新規メタ開裂酵素であることが示唆された。 -ketoadipateに、HQは-hydroxymuconic semialdehydeにそれぞれ変換されることが判明した。これにより、LinEは既知のカテコール型の基質(1,2位に水酸基がついている)を開裂するメタ開裂酵素とは異なり、ハイドロキノン型の基質(1,4位に水酸基がついている)を開裂する新規メタ開裂酵素であることが示唆された。 4.LinEの精製とその特徴の解析 LinEの性質を調べるため、LinEを大量発現させた大腸菌の粗酵素抽出液をDEAE・TOYOPEARL、BUTYL・TOYOPEARL、ゲル濾過の各種クロマトグラフィーに供し、SDS・PAGE上で一本のバンドになるまで精製した。酸素電極を用いて基質添加時の酸素消費量を測定した結果、基質としてCHQ、HQを加えた際には酸素を消費するが、カテコールを基質としたときは酸素を消費しないことが明らかとなった。反対に、コントロールとして用いた代表的なメタ開裂酵素であるXylE(catechol 2,3-dioygenase)はカテコールを加えたときにのみ酸素を消費する。この結果より、LinEはハイドロキノン骨格を持つ基質に対して活性を持つ、新規メタ開裂酵素であることが確認された。 5.linD、linEの発現制御機構及びこれらを含むオペロンの解析 構成的に弱いLinD活性を保持する変異株UT102の変異部位を同定したところ、変異部位はlinEの上流にあり、UT26中のこの部位にはlinEと逆向きに909bpのORFが存在した。相同性検索の結果、このORFはLysRファミリーに属する転写調節因子と相同性があったため、このORFをlinRと命名した。linEの上流にはLysRファミリータンパクの認識配列であるパリンドームを含むTN11Aの配列が存在する。この配列を含むlinEの上流配列及びこれに続くlinEのN末をlacZにin frameつないだプラスミド中にラクトースプロモーター下につないだlinRを挿入したプラスミドを作製し、これを保持する大腸菌に-HCH、2,5-DCHQ、CHQ、HQをそれぞれ与えてそれぞれのLacZ活性を測定した。その結果、2,5-DCHQ、CHQを与えたときにLacZ活性が上昇した。また、UT26、及びlinR変異株UT102に対して2,5-DCHQ、CHQ、HQの存在下でlinEをプローブとしたノーザン解析を行った結果、UT26中ではこれらの基質の存在下でlinEの発現は上昇するが、UT102ではlinEの発現は抑えられていることが判明した。以上の結果より、LinRはLinD、LinEの基質の存在下でlinD、linEの転写を活性化する転写活性化因子であることが示された。また、linD、linEの近傍には、これらと共通の発現制御を受けていると思われる4つのORFが見出されたので、これらの機能についても解析を行った。 6.まとめ 本研究では、UT26における-HCH分解代謝に関与する酵素遺伝子としてlinD、linE、linRを単離し、その解析を行った。その結果、UT26における-HCHの分解代謝経路は下図の様になった。微生物による芳香族の分解代謝経路の多くは、そのキーステップとなる環開裂の基質としてカテコール型の物質を用いる。一方、UT26の-HCH分解経路は、hydroquinone型の(chloro)hydroquinoneを環開裂の基質とする、現在までに知見の少ない経路である。hydroquinoneを直接開裂する反応は現在までにいくつか報告されているが、それに関与する酵素遺伝子の単離は本研究のlinEが始めてであり、linEと一般的なメタ開裂酵素との立体構造の違いに興味が持たれる。また、linA、linB、linCがオペロンを構成しておらず構成的に発現しているのに対して、linD、linEはオペロンを形成し、その発現は転写因子LinRによって制御されている。このことは、UT26が-HCH分解代謝能を獲得するに当たって、上流の代謝に関与する酵素遺伝子を外部から獲得し、それと共に以前から保持していた(比較的発達した制御機構を持つ)下流の代謝経路を進化させた可能性を示唆しており、非常に興味深い。さらに下流の代謝経路、及びそれに関与する酵素遺伝子を同定することによって本菌の-HCH代謝能獲得機構についての新たな知見が得られることが期待される。  | 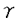 -HCH分解系に関する分子遺伝学的研究
-HCH分解系に関する分子遺伝学的研究