蛋白質の燐酸化・脱燐酸化の現象は、医学的にも生物学的にも重要な意義を持つことが明らかにされている。現在、ほとんど全ての細胞の制御について蛋白質の燐酸化が関与していることが示唆されており、従って蛋白質燐酸化あるいは脱燐酸化の過程を解明することは、様々な生命現象を理解する上で不可欠であると考えられる。 蛋白質の燐酸化の過程は、燐酸化を触媒するプロテインキナーゼ(protein kinase;PK)と脱燐酸化を触媒するプロテインフォスファターゼ(protein phosphatase;PP)によって、可逆的に制御されている。このような蛋白質の燐酸化は、その蛋白質のもつ活性を別の段階に切り替えるスイッチのような役割を果たしていると考えられている。原癌遺伝子として最初に発見された遺伝子c-srcがPTK(protein tyrosine kinase)であることが見いだされて以来、蛋白質のチロシン燐酸化が細胞の癌化や分化に重要であることが注目されてきた。今ではEGF(epidermal growth factor)やFGF(fibroblast growth factor)など増殖因子の受容体の多くがPTKであることが報告されている。また、NGF(nerve growth factor)、BDNF(brain-derived neurotrophic factor)、NT-3(neurotrophin 3)などの神経細胞に増殖と分化を誘導する分子の受容体もPTKであることが解り、神経系におけるチロシン燐酸化の重要性が示唆されている。 これまでにPTKによる蛋白質の燐酸化については数多く報告されてきたが、PTP(protein tyrosine phosphatase)による蛋白質の脱燐酸化については報告が少ない。これは各々のPTPに対する特異的な基質や阻害剤が不明なため、個々のPTPの脱燐酸化反応を評価できるような実験が困難なことに多く起因している。従って、PTPの細胞内における機能についてはいまだ不明な点が多いのが現状である。前述したように蛋白質の燐酸化と脱燐酸化は可逆的に調節されており、PTKだけでなくPTPの側から解析することは不可欠である。筆者は、このようなPTPの機能研究における実験手技的な問題点を避け、発生工学の手法を用いてPTPの生体内における機能を明らかにすることを試みた。 筆者はチロシン特異的なフォスファターゼであるMPTP (murine protein tyrosine phosphatase )遺伝子に注目し、Cappechiらによって確立された標的遺伝子組換(gene targeting)を用いてMPTP遺伝子に変異を導入したマウスの表現型を解析した。ある特定な遺伝子産物の機能を探索する際、生体内でその遺伝子を過剰発現させたり、あるいは機能欠損させることによって得られる表現型を解析することは、有用な方法である。特に免疫系や中枢神経系などのように多種多様な細胞、組織が複雑に関わるような生命現象については重要な解析方法である。 (murine protein tyrosine phosphatase )遺伝子に注目し、Cappechiらによって確立された標的遺伝子組換(gene targeting)を用いてMPTP遺伝子に変異を導入したマウスの表現型を解析した。ある特定な遺伝子産物の機能を探索する際、生体内でその遺伝子を過剰発現させたり、あるいは機能欠損させることによって得られる表現型を解析することは、有用な方法である。特に免疫系や中枢神経系などのように多種多様な細胞、組織が複雑に関わるような生命現象については重要な解析方法である。 MPTP遺伝子はフォスファターゼドメインの保存された領域をプローブとして用いたクロスハイブリダイゼーション法によって水野博士らにより単離された。MPTPは、細胞質内に酵素活性ドメインを2つ持ち、細胞膜貫通領域、さらに細胞外領域を持つ受容体型チロシンフォスファターゼである。細胞外領域はN-末端側のイムノグロブリン(Ig)様ドメインの繰り返し構造と、C-末端側のフィブロネクチンタイプIII(FN III)ドメインの繰り返し構造によって特徴づけられる。MPTPのリガンドや基質については全く不明であり、細胞内における機能についてはほとんど解析されていない。しかし、これらの分子の細胞外領域の構造的な特徴は、細胞接着に関与する分子群の構造と類似しており、細胞接着の情報を細胞内に伝えるシグナル伝達因子として重要である可能性が示唆され、興味深い。 筆者はMPTP遺伝子の酵素活性に必須な部分を欠損させた変異マウスの作製に成功し、その表現型について解析した。その結果、MPTP遺伝子変異マウスは成長が遅く、離乳期までの死亡率が非常に高いことが解った(図1)。また、このような成長遅延と高い死亡率を示す原因として、餌の摂取に何らかの問題があり栄養不良になることが示唆された。MPTP遺伝子は胸腺や脾臓で部位特異的に発現しており、免疫系臓器において機能的役割が期待されていた。しかし胸腺や脾臓の各免疫細胞の割合は正常で、マイトジェンに対する応答性に差はなく、組織学的にも正常であると思われた。  図1 MPTP遺伝子変異マウスの体重曲線と生存率エサが上部(top)にる場合のMPTP(-/-)(白丸)とエサを床面(floor)に置いた場合のMPTP(-/-)(黒丸)と野生型(+/+)(黒四角)マウスの生存率(左図)と体重曲線(右図)を示した。エサをケージ上部(top)に置く通常の飼育条件下では、MPTP(-/-)は生後35日までに60%が死亡するが、エサをケージ床面(floor)に置くだけで生存率が上がる(黒丸)(左図)。しかし、野生型(+/+)と比較すると体重が軽く矮小であることがわかる(右図)。 図1 MPTP遺伝子変異マウスの体重曲線と生存率エサが上部(top)にる場合のMPTP(-/-)(白丸)とエサを床面(floor)に置いた場合のMPTP(-/-)(黒丸)と野生型(+/+)(黒四角)マウスの生存率(左図)と体重曲線(右図)を示した。エサをケージ上部(top)に置く通常の飼育条件下では、MPTP(-/-)は生後35日までに60%が死亡するが、エサをケージ床面(floor)に置くだけで生存率が上がる(黒丸)(左図)。しかし、野生型(+/+)と比較すると体重が軽く矮小であることがわかる(右図)。 興味深いことにMPTP遺伝子変異マウスは行動学的に自発的運動量の増加や運動障害を示し、さらにモリス水迷路(図2)・八方向迷路・T迷路試験で学習能力が低下していることが明らかとなった(エーザイの小倉博士らとの共同研究)。このような行動の異常から、MPTPが中枢神経系で重要な働きを担うことが強く示唆された。MPTPと非常に相同性の高いDLAR遺伝子に変異を持つショウジョウバエが、運動神経の軸索走行の異常を示した報告からも、MPTP遺伝子変異マウスが運動障害の表現型を示したことは非常に興味深い。 MPTP遺伝子は中枢神経系において、部位特異的な発現が報告されている。MPTP遺伝子は、海馬のCA2からCA3のごく限られた領域の神経細胞で強く発現しており、他に視床網様核および皮質梨状葉の神経細胞で発現している。さらに、ラットの相同遺伝子であるRPTPは胎生期の中枢神経系において、大脳皮質、間脳、脳下垂体、内耳の神経細胞および脊髄運動神経に特異的に発現していることが報告され、神経細胞の発達分化あるいは細胞機能に関与することが示唆されていた。MPTP遺伝子変異マウスの脳を組織学的に解析した結果、明らかな差が無かったことから、筆者はMPTPは脳の構造構築に関わるのではなくより高次な機能に関わると考え、海馬の神経シナプスの性状について電気生理学的に解析した(ERATOの加藤博士らとの共同研究)。  図2 モリス水迷路試験(プローブ試験)7日間到達台の位置を学習させた後に到達台を取り除き、1分間マウスを泳がせ到達台のあった区画(SE)を泳いでいた時間を計測した。野生型マウス(+/+)はSE区画を泳いでいた時間が有意に高かったが(ANOVA検定;・p〓0.001)、変異型マウスでは差がなかった(ANOVA検定;p=0.116) 図2 モリス水迷路試験(プローブ試験)7日間到達台の位置を学習させた後に到達台を取り除き、1分間マウスを泳がせ到達台のあった区画(SE)を泳いでいた時間を計測した。野生型マウス(+/+)はSE区画を泳いでいた時間が有意に高かったが(ANOVA検定;・p〓0.001)、変異型マウスでは差がなかった(ANOVA検定;p=0.116) 現在、シナプス可塑性(シナプス伝達効率の永続的変化)の実験モデルとして、海馬のLTP(long-term potentiation)が提唱されている。LTPとはシナプスの強さ(シナプス前の刺激に対するシナプス後の応答)の長期的な増強である。先に述べたようにMPTP遺伝子変異マウスは学習機能に障害があり、驚いたことに海馬のLTPがより増強されていることが解った(図3)。従ってMPTP遺伝子変異マウスの学習能力の低下は、海馬のLTPの増強というシナプス可塑性の変化に起因することが示唆された。非特異的PTP阻害剤を用いた実験からPTPがLTPに関わることが示唆されていたが、本研究によりそのPTPのひとつが初めて明らかにされた。これまでに海馬LTPの制御メカニズムとして、NMDA型グルタミン酸受容体のPTKを介するチロシン燐酸化が注目されている。NMDA型グルタミン酸受容体の活性化はLTPに必須であると考えられており、このような過程にMPTPは関与するのかもしれない。また、細胞接着因子であるカドヘリン分子がLTPに関与することが報告されているが、その下流にやはりチロシン燐酸化が関わることが示唆されておりMPTPとの関連が興味深い。  図3 海馬の電気生理学的解析海馬のスライス標本を用い、テタヌス刺激後の(A)CA1領域、(B)CA3領域のLTPを記録した。いずれの領域でも野生型(黒丸)に較べ、MPTP変異型(白丸)マウスで有意にLTPが増強されていた。右に図中a点とb点の代表的な波形を示した。 図3 海馬の電気生理学的解析海馬のスライス標本を用い、テタヌス刺激後の(A)CA1領域、(B)CA3領域のLTPを記録した。いずれの領域でも野生型(黒丸)に較べ、MPTP変異型(白丸)マウスで有意にLTPが増強されていた。右に図中a点とb点の代表的な波形を示した。 さらに、海馬LTPの低下により学習能力が低下することは良く知られているが、海馬LTPの増強と学習能力の低下を同時に示した変異マウスの報告は世界でも例が少なく、学習と海馬LTPの関係を研究する上で非常に重要な発見といえる。筆者の研究は、学習や記憶のような神経回路を形成するためには、適切な範囲で起こるLTP(シナプス可塑性)が重要であるという仮説を支持する結果となった。 以上のように、筆者はMPTP遺伝子変異マウスを作製することにより、MPTPが中枢神経系で重要な働きを担うことを示す解析結果を得、同時に学習や記憶といった高次脳機能について新たな知見を得たので報告する。 | 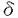 の機能解析
の機能解析