心筋細胞の収縮・弛緩は細胞内Ca2+濃度([Ca2+]i)の増減によって制御される。心筋の[Ca2+]iの増加において、細胞内Ca2+貯蔵部位である筋小胞体(SR)からのCa2+放出(Ca2+-induced Ca2+ release;CICR)が重要な役割を果たすことが示されているが、[Ca2+]iの増加に対するCICRの関与には動物種差があることも報告されており、統一的な見解には至っていない。また、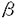 受容体刺激は収縮蛋白質のCa2+感受性を減少させることが報告されているが、充分には検討されていない。本研究は、モルモットの右心室乳頭筋において、単収縮はCICRとCa2+流入のどちらに強く依存しているのか、そして、受容体刺激による収縮蛋白質Ca2+感受性減少がどの程度収縮張力の制御に関与するのかを明らかとすることを目的として行われた。 受容体刺激は収縮蛋白質のCa2+感受性を減少させることが報告されているが、充分には検討されていない。本研究は、モルモットの右心室乳頭筋において、単収縮はCICRとCa2+流入のどちらに強く依存しているのか、そして、受容体刺激による収縮蛋白質Ca2+感受性減少がどの程度収縮張力の制御に関与するのかを明らかとすることを目的として行われた。 1.モルモット心筋における[Ca2+]iと単収縮の関係 外液Ca2+濃度の増加によって[Ca2+]i、収縮の両者は増加した。刺激頻度の増加によっても両者は増加するものの、[Ca2+]iの上昇は比較的小さなものであった。同様の傾向は強心配糖体であるウワバインや電位依存性Ca2+チャネル活性化薬であるBay K8644の投与時にも観察された。一方、[Ca2+]iの上昇速度は収縮の増加に比例した上昇を示した。 CICRを抑制する薬物であるライアノジンにより、[Ca2+]iはおよそ30%抑制された。ラットにおいては、ライアノジンにより[Ca2+]iが60-90%抑制されることが知られており、モルモットでは[Ca2+]iの上昇に占めるCICRの割合がラットより低いことが示唆された。さらに、[Ca2+]iの上昇速度の増加はライアノジンの投与によってほぼ完全に抑制された。 [Ca2+]iと収縮張力、[Ca2+]iの上昇速度と収縮張力は、両者とも高い相関を示したが、後者のほうがより高い相関を示した。一方、ライアノジン存在下では、いずれの場合も相関開係の著しい減弱が見られた。 2.受容体刺激の影響 モルモットにおいて、単収縮の弛緩相における[Ca2+]i-収縮の関係に対して、受容体作動薬であるイソプロテレノールは影響を及ぼさなかった。さらに、強縮後の弛緩相を用いて定常状態での[Ca2+]iと収縮の関係も検討した。ラット心室筋においては、24℃ではCa2+感受性の減少が観察されたものの、30℃では観察されなかった。モルモット心室筋では、24℃、30℃のいずれの温度においても、イソプロテレノール、アデニル酸シクラーゼ活性化薬のフォルスコリン、の両薬物によってCa2+感受性は変わらなかった。 モルモット乳頭筋の -toxinによる脱膜化標本において、cAMP依存性蛋白質リン酸化解素(protein kinase A:PK-A)を活性化するcAMPの投与は、SRのCa2+取り込みを増強したが、収縮のCa2+感受性には影響を及ぼさなかった。一方、サポニンによる脱膜化標本では、過剰量のPK-AのcAMPとの同時投与により、Ca2+感受性は減少した。 -toxinによる脱膜化標本において、cAMP依存性蛋白質リン酸化解素(protein kinase A:PK-A)を活性化するcAMPの投与は、SRのCa2+取り込みを増強したが、収縮のCa2+感受性には影響を及ぼさなかった。一方、サポニンによる脱膜化標本では、過剰量のPK-AのcAMPとの同時投与により、Ca2+感受性は減少した。 モルモット右心室乳頭筋において、イソプロテレノールは、単収縮と[Ca2+]iを増加させ、[Ca2+]iと収縮張力は2相性の変化を示した。1相目はライアノジンにより消失したことからCICRに依存し、ライアノジンに影響を受けなかった2相目はCa2+流入を介するものと考えられた。 以上の成績から、モルモット右心室乳頭筋においては、[Ca2+]iの上昇速度はCICRを反映すること、また、[Ca2+]i上昇に占めるCICRの割合は低いにも関わらず、CICRは単収縮の大きさの重要な決定因子であることが明らかとなった。このように、[Ca2+]i増加機構ごとに収縮発生の有効性が異なるのは、細胞内での[Ca2+]iの局在が一因であると考えられた。さらに、受客体刺激による収縮蛋白質のCa2+感受性の減少には種差があり、加えて、温度依存性にその作用が減弱することから、生理的な条件下では受容体刺激によるCa2+感受性の減少は発現しておらず、細胞内Ca2+動態の修飾が、受容体刺激の主な作用であることが明らかとなった。これらの知見は学術上の重要性はいうに及ばず、心臓作動性の医薬品の開発にとっても有用な知見と考えられる。よって、審査委員一同は本論文が博士(獣医学)の論文として価値あるものと認めた。 |