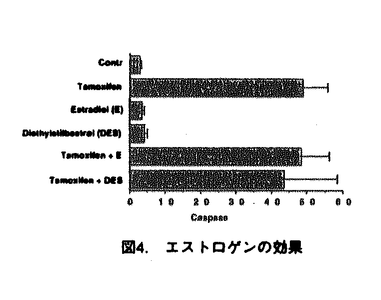
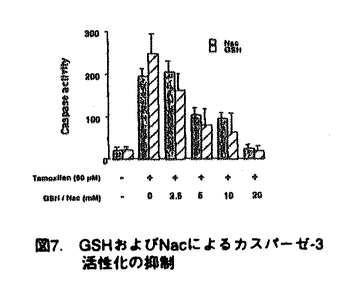
本研究は、乳癌のホルモン療法に使用されている抗エストロゲン剤であるタモキシフェンの、副作用として報告される白血球の減少が、エストロゲンレセプター(ER)非依存的なアポトーシス誘導による可能性を明らかにするため、ERを持たない白血病細胞U-937を用いて、タモキシフェンのアポトーシス誘導能、およびその作用経路について解析を試みたものであり、下記の結果を得ている。 1.タモキシフェンをU-937細胞に添加すると、濃度および時間依存的に生細胞数が減少し、この細胞死がアポトーシスであることを、アポトーシスに特徴的な断片化DNAの検出により確認した。さらに、アポトーシスにおいて特徴的に活性化されるプロテアーゼであるカスパーゼの活性化を、特異的基質を用いた活性測定、および活性化カスパーゼ-3特異的抗体を用いたイムノブロット法で調べ、カスパーゼ-3の活性化を確認した。カスパーゼ-3の上流で作用するとされるカスパーゼ-1の活性は検出されなかったことから、タモキシフェンによるアポトーシス誘導においては、カスパーゼ-3の活性化は、カスパーゼ-1が関与しない経路によることが示された。 2.エストロゲン自身ではアポトーシスは誘導されず、またタモキシフェン添加時にエストロゲンが共存しても競合阻害が認められないことから、タモキシフェンによるアポトーシス誘導は、ER非依存性の作用機所であることが示された。また、RNA/タンパク質合成の阻害剤の存在下でもアポトーシスが誘導されたことから、タモキシフェンによるアポトーシス誘導には、タンパク質の新規合成は不必要であることが示された。 3.タモキシフェン添加15分後より、ミトコンドリアから細胞質内ヘシトクロムcが流出することが、イムノブロット法で示された。さらに、ミトコンドリアの膜電位の変化を調べ、タモキシフェン添加後の膜電位の低下が示された。また、細胞内の還元型グルタチオン(GSH)濃度の変化を測定し、タモキシフェン添加後、GSH濃度が急速に減少することが示された。しかしながら、タモキシフェン添加後の活性酸素種の発生は検出されなかったことから、GSHの減少は細胞内の酸化還元状態の変化以外に、GSHが細胞外へ排出される可能性が考えられた。 4.タモキシフェンによるアポトーシス誘導時に、GSHあるいはN-アセチルシステインを添加することにより、タモキシフェンによるカスパーゼの活性化、DNAの断片化、およびシトクロムcの細胞質への流出が抑制されることが示された。GSHの減少はタモキシフェン誘導アポトーシスのキーイベントである可能性が考えられた。また、タモキシフェンの作用経路はミトコンドリアの膜における機能的あるいは物理的な変化であると考えられた。 以上、本論文はU-937細胞において、タモキシフェンによるアポトーシスの誘導機構を解析し、その作用機所がエストロゲンレセプター非依存的で、直接的あるいは間接的にミトコンドリアに作用することを明らかにした。 本研究はこれまで知見のなかった、タモキシフェンによる細胞毒性のメカニズムの解明に重要な貢献をなすものと考えられ、学位の授与に値するものと考えられる。 |