1.はじめに JCウイルス(JCV)は1971年Padgettらによって、進行性多巣性白質脳症(PML)患者の脳からはじめて分離された。PMLは免疫機能の低下した患者にまれに起きる疾患であるが、近年AIDS患者の増加に伴い発症例が増えている。他方、JCVはヒト集団に広く浸淫しているウイルスでもある。JCVの初感染は幼児期に起き、無症候性である。初感染の後、JCVは腎に寄生する。ほとんどの成人は仔ウイルスを尿中に排出するので、JCV DNAはPCRにより容易に分離できる。
JCVの伝播の特徴は、水平感染にも関わらず、主に親から子へ伝播することである。また、異なるヒト集団間ではJCVはほとんど伝播しないことも知られている。これらのことから、JCVは日常的に密接な接触のある限られた集団(通常は家族)の中で伝播することが明らかである。
これまでに旧世界(ヨーロッパ、アフリカ、アジア)の各地のJCV DNAが制限酵素を用いて解析され、JCV DNAが3つの型(A、B、C)に分類されることが明らかにされた。そして、それぞれの型は特徴的な地理的分布を示すことが発見された。すなわち、Aはヨーロッパ、Bはアジアとアフリカ、Cはアフリカの一部に分布した。このことからJCVゲノム型とヒト集団と間には密接な関連があることが示唆された。
本研究では先ず、旧世界各地で採取された尿から得たJCV DNAを分子系統解析することによって、旧世界のJCV DNAをゲノム型に分類し、各ゲノム型の地理的分布を詳細に明らかにすることを目的とした。次ぎに、この結果を様々な民族の起源を探る上でのカタログとし、その例として中南米先住民と東シベリア先住民の起源を推定することを試みた。
2.旧世界におけるJCVゲノム型の分類とその分布 旧世界(ヨーロッパ、アフリカ、アジア)の34カ国、42地域で尿を採取した。遠心分画した尿からウイルス粒子を回収し、ウイルスDNAを抽出した。これを鋳型として、610塩基対のVT遺伝子間領域(IG領域と略す。この領域が最も塩基置換が多い)を増幅した。得られた増幅断片をpUC19にクローニングして、オートシークエンサーによって塩基配列を決定した。これらの塩基配列から近隣結合法により分子系統樹を作成した。
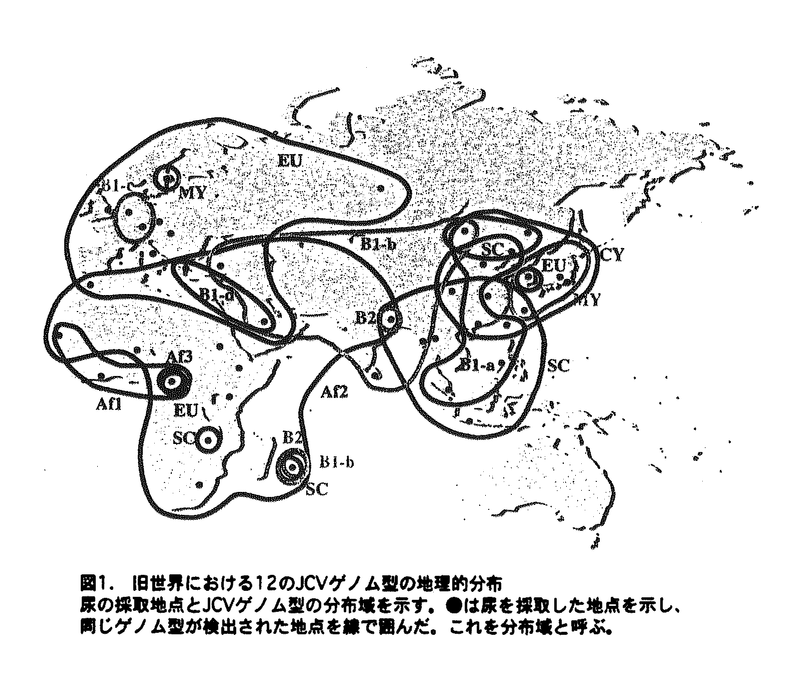
分子系統樹から、旧世界のJCVは12のゲノム型に分類できた。これらのゲノム型の旧世界における分布域を図1に示した。各ゲノム型の分布域を要約する。EU(ヨーロッパ全域、北アフリカ、西アジア)、Af1(西アフリカの一部)、Af2(アフリカ全域、中東、インド北部)、Af3(中央アフリカ)、B1-a(中国、フィリピン)、B1-b(西アジア、中央アジア)、B1-c(ヨーロッパ)、B1-d(中東)、B2(インド、モーリシャス)、SC(東南アジア、中国南部)、CY(北東アジア)、MY(日本、韓国)。
6つのゲノム型(EU,Af2,B1-a,B1-b,SC,CY)は広域に分布していた。残り6つのゲノム型(Af1,Af3,MY,B1-c,B1-d,B2)は比較的狭い分布域を有し、広大な他のゲノム型の分布域の中に存在した。
JCVゲノム型の分布域はそれぞれ特徴的であったことから、地球上の様々なヒト集団がそれぞれ固有のJCVゲノム型を持つことが示唆された。また、JCVゲノム型の分布パターンは、アフリカで誕生した人類のその後の分化および移動と一致した。JCVゲノム型ほどきれいにヒト集団との関連が示された遺伝マーカーは知られていない。以上から、JCVゲノム型解析により、ヒト集団の移動の軌跡をたどることが可能であることがわかった。
JCVゲノム型解析のもうひとつの利点は、主要なゲノム型の分布域の中に少数ゲノム型を検出できることである。このようにヒト集団の祖先のわずかな痕跡も見つけだすことができる。
3.JCVから見た先住民の起源3-1.中南米先住民 これまでの様々な人類学的、考古学的研究から、アメリカ先住民の祖先は1.5〜3万年前にベリンジアを渡ってきたモンゴロイドであると考えられている。しかしアメリカ先住民が現存するモンゴロイド集団のうちのどの集団に一番近いかに関しては諸説がある。
グアテマラ(3地域)とペルー(2地域)において、各地域約50人の先住民から尿を採取し、IG領域を増幅した。得られたIG領域の塩基配列と旧世界の塩基配列から、近隣結合法により分子系統樹を作成した。
グアテマラとペルーの先住民が保有するJCVはMYに属することが明らかになった。MYは旧世界では主として日本と韓国に分布するゲノム型である。また、系統樹解析からMYはさらに3系統(MY-a、MY-b、MY-c)に分かれることが見いだされた。MY-aは主に日本と韓国の株、MY-bは主にグアテマラの株、MY-cは主にペルーの株を含んでいた。
本研究の結果から、中南米先住民は日本人・韓国人と近縁のモンゴロイドであることが示唆された。1〜3万年前にベリンジアで広く活動していたモンゴロイド集団のひとつがMYを保有していたと推定される。現在のMYの地理的な分布はこの集団のその後の移動を示していると考えられる。
3-2.東シベリア先住民 シベリアには多数の先住民族が入り組んで分布しており、その起源は必ずしも明確にはされていない。また、シベリア先住民はシベリアの人口の90%以上を占めるヨーロッパ系移民との混血も進んでいる。このような集団からも、JCVゲノム型解析では先住民固有のゲノム型を見つけだすことが期待できる。
ヤクーツクのヤクート族、アムール川流域のナーナイ族およびコリマ川流域の複数集団から尿を採取した。また、対照としてハバロフスクのスラブ系住民からも尿を採取した。得られたIG領域の塩基配列と旧世界の塩基配列から、近隣結合法により分子系統樹を作成した。
(1)ヤクート族の主要なゲノム型はB1-bであった。B1-bは西アジアと中央アジアに存在するゲノム型である。(2)ナーナイ族とコリマ川流域先住民の主要なゲノム型はEUであった。EUは主にヨーロッパに存在する。しかし、シベリアのEUの多くは系統樹の上でヨーロッパのものとは異なる独自のクラスター(EU-a)を形成した。(3)ハバロフスクのスラブ系住民の主要なゲノム型はEUで、これはヨーロッパのEUと同じクラスターに分類された。
以上のことから、ヤクート族の祖先は西アジア・中央アジア系の集団であることが示唆された。これはヤクート族の人類学的な分類と一致する。一方、ナーナイ族とコリマ川流域先住民の祖先集団の一部がコーカソイド系であることが示唆された。これは従来の人類学的な研究では得られなかった新しい知見である。