インターロイキン-2(IL-2)は、リンパ球の増殖及び分化を誘導することにより、免疫反応に重要な役割を果たしている。機能的高親和性IL-2受容体(IL-2R)は、IL-2R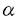 、IL-2R 、IL-2R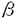 c、IL-2R c、IL-2R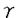 c鎖の3つのサブユニットから構成されている。IL-2Rc鎖はIL-15Rにも共有され、IL-2Rc鎖はIL-4R、IL-7R、IL-9R、IL-15Rにも共有されるサブユニットである。 c鎖の3つのサブユニットから構成されている。IL-2Rc鎖はIL-15Rにも共有され、IL-2Rc鎖はIL-4R、IL-7R、IL-9R、IL-15Rにも共有されるサブユニットである。 IL-2Rc鎖及びIL-2Rc鎖の細胞内領域は、各種シグナル伝達分子の活性化に重要な役割を果たしている。IL-2Rc鎖の細胞内領域は、セリン残基に富むS領域、酸性アミノ酸に富むA領域、プロリン残基に富むH領域とから構成されている。膜近傍のS領域は、Jak1蛋白チロシンキナーゼ(PTK)及びSykPTKとの会合に必要であり、S領域に隣接するA領域は、src-ファミリーPTKLckやShcとの会合に必要である。S領域、A領域はまた、プロトオンコジーンの転写誘導にも必須である。一方、C末端のH領域は、IL-2刺激によるStat5の活性化に必須である。 IL-2Rc鎖の細胞内領域は86アミノ酸から構成され、Jak1、Jak3、及びStat5の活性化、c-fos、c-myc、bcl-2の転写誘導に必要な領域である。なかでも、Jak3は、IL-2依存性細胞増殖に重要な役割を担っている。Jak3は、IL-2刺激によるc-fos、c-mycの転写誘導に必須であり、また、転写因子Stat3及びStat5を活性化する。IL-2Rc鎖の細胞内領域のうちC末端48アミノ酸が、Jak3との会合及びその活性化に関与しているが、Jak3との会合に関与しないIL-2Rc鎖の膜近傍領域の役割は、いまだ明らかではない。 本研究では、IL-2シグナル伝達経路の活性化における、IL-2Rc鎖細胞内領域の膜近傍部と膜遠位部の役割を、培養細胞系と発生工学的手法を用いて解明した。最初に,IL-2Rc鎖の野生型及び変異体のcDNAを導入した培養細胞系を用いて,C末端48アミノ酸を欠失したc鎖変異体が、Jak3の活性化なしに、Jak1及びStatの活性化を含むシグナルを伝達しうることを示した。これらの現象は、IL-2Rc鎖膜近傍領域のプロリンに富むモチーフに点変異を導入することにより消失した。これらの結果は、IL-2Rc鎖の膜近傍領域によって誘導されるJak3非依存性経路の存在を示唆している。しかし、培養細胞系での解析では、この経路がリンパ球系の分化増殖においていかなる役割を担っているかは解明できない。そこで、発生工学的手法により、IL-2Rc鎖の野生型及び変異体のトランスジーンをIL-2Rc鎖欠損マウスのリンパ球に発現させた。興味深いことに、C末端48アミノ酸を欠失したc鎖変異体を発現させることによって、胸腺細胞数,末梢T細胞数が、IL-2Rc鎖欠損マウスに比し回復し、IL-2Rc鎖の膜近傍領域からのシグナル伝達経路がTリンパ球の分化に重要な役割を担っていることが示唆された。さらに、このJak3非依存性シグナル伝達経路にbcl-2の転写誘導が関与している可能性が示された。一方、発生工学的手法で得られた結果はまた、IL-2以外の刺激によりJak3を活性化させる経路の存在をも示唆していた。 第一部 培養細胞系を用いたIL-2Rc鎖の機能解析 ヒトIL-2Rc鎖の野生型、細胞内領域のC末端79アミノ酸を欠失するM1変異体、C末端48アミノ酸を欠失するM2変異体のcDNAを、IL-2Rc鎖を欠くヒトT細胞株ED40515(-)細胞(以下ED細胞)に導入し、JakPTKのチロシンリン酸化とStatの活性化を検討した。 野生型IL-2Rc鎖をED細胞株に発現させたWT-24細胞では、IL-2刺激にてJak1及びJak3の活性化が認められたが、M1変異体を発現させたM1-4細胞では活性化は消失していた。一方、M2変異体を発現させたM2-7細胞では、Jak3の活性化は完全に消失していたが、Jak1の活性化は認められた。この結果から、IL-2Rc鎖の膜近傍38アミノ酸からなる領域が、Jak3の活性化なしにJak1の活性化に至るシグナルを伝達しうることが示された。また、WT-24細胞で認められた、IL-2刺激によるStat3及びStat5の活性化は、M1-4細胞では消失したが、M2-7細胞では認められた。さらに、IL-2刺激による phosphatidylinositol 3-kinase(PI3K)のp85サブユニットのチロシンリン酸化が、WT-24細胞のみならずM2-7細胞でも認められた。これらの結果は、IL-2Rc鎖の膜近傍領域が、Jak3の活性化なしにStatの活性化及びPI3Kのp85サブユニットのリン酸化を誘導しうることを示し、これらのシグナル伝達経路に、Jak3非依存性にJak1の活性化が関与している可能性を示唆する。但し、M2-7細胞におけるJak1のリン酸化、Statの活性化及びp85サブユニットのリン酸化の程度は、いずれもWT-24細胞に比し有意に低く、Jak3が、Jak1の活性化を増強することにより、IL-2シグナル伝達経路を増強することが示唆された。 一方、IL-2刺激によるmitogen-activated protein kinase(MAPK)の活性化は、WT-24細胞では認められたが、M2-7細胞では認められず、MAPK活性化が、IL-2Rc鎖の膜遠位領域を介したシグナル伝達経路に全面的に依存していることが示された。 本研究ではさらに、IL-2Rc鎖の膜近傍領域によるシグナル伝達の分子生物学的機序に迫るために、他のサイトカイン受容体サブユニットにも共通に見出される、膜近傍領域のプロリンに富むモチーフに着目した。IL-2Rc鎖M2変異体のこの領域の2つのプロリン残基をアラニンで置換したPA変異体を発現するED細胞株(PA-2細胞)を樹立した。M2-7細胞で認められた、IL-2刺激によるJak1、Statの活性化、及びPI3Kのp85サブユニットのチロシンリン酸化は、PA-2細胞では完全に消失した。これらの結果から、このプロリンに富むモチーフが、IL-2Rc鎖の膜近傍領域を介したJak3非依存性シグナル伝達経路に重要な役割を担っていることが示唆される。 第二部 発生工学的手法を用いたIL-2Rc鎖の機能解析 より生理的に意義のある条件下でのIL-2Rc鎖の細胞内領域の機能を解明することを目的として、発生工学的手法による機能解析を行った。マウスIL-2Rc鎖の野生型及びC末端48アミノ酸を欠失するM2変異体のcDNAを、ヒトCD2プロモーター制御下にリンパ球特異的に発現させたトランスジェニックマウスを作製し、IL-2Rc鎖欠損マウスとの交配を繰り返すことにより、IL-2受容体の発現を再構築したマウスを作製した。 IL-2Rc鎖欠損マウスでは、胸腺細胞、及び末梢T細胞、B細胞が野生型に比し著減しているが、M2変異体を発現させたcM2:-/Yマウスでは、胸腺細胞数はc鎖欠損マウスに比し著明に回復、脾T細胞数、B細胞数もc鎖欠損マウスに比し回復した。この結果は、IL-2Rc鎖の膜近傍領域が、T細胞、B細胞の分化に必須のシグナルを伝達することを示している。 Jak3欠損マウスでは、IL-2Rc鎖の膜近傍領域からのシグナルは伝達されるにもかかわらず、胸腺細胞数、末梢T細胞数、B細胞数はIL-2Rc鎖欠損マウスと同様に著減したままであることが報告されている。しかし、興味深いことに、cM2:-/Yマウスでは、IL-2Rc鎖を介したJak3の活性化が起こり得ないにもかかわらず、T細胞の分化は回復した。このことは、IL-2Rc鎖とは独立に、Jak3を活性化するシグナル伝達経路が存在する可能性を示唆する。 さらに、IL-2Rc鎖の膜近傍領域がT細胞の分化に作用する機序として、bcl-2の活性化に着目した。実際、cM2:-/Yマウスでは、CD4陽性脾細胞におけるbcl-2の発現レベルが、c鎖欠損マウスに比し有意に高く、IL-2Rc鎖の膜近傍領域を介したT細胞の分化誘導に、bcl-2の転写を誘導する経路が関与している可能性が示唆された。 一方、cM2:-/Yマウスでは、T細胞の分化能は回復するにもかかわらず、ConA刺激、抗CD3 抗体刺激による細胞増殖は認められず、末梢T細胞の増殖にはJak3の活性化が必須であることが示された。 抗体刺激による細胞増殖は認められず、末梢T細胞の増殖にはJak3の活性化が必須であることが示された。 以上の一連の結果は、IL-2Rc鎖からのJak3非依存性経路の存在を示し、この経路がリンパ球の分化に関与していることを示すとともに、Jak3をIL-2Rc鎖非依存性に活性化する未知の経路の存在を強く示唆しており、リンパ球の分化増殖に関与する細胞内シグナル伝達経路の研究において新たな知見を提供するものである。 |  c鎖の細胞内領域のin vitro、in vivoにおける機能解析
c鎖の細胞内領域のin vitro、in vivoにおける機能解析