| [研究の背景および目的] Helicobacter pyloriはヒト胃粘膜に感染するグラム陰性桿菌であり、潰瘍の発症においては重要な役割を果たしているものと考えられている。特に、H.pyloriの除菌によって潰瘍の再発が抑制されることは潰瘍との関連を強調する事実である。また、近年胃癌の発生との関連が注目されてきている。しかし、H.pyloriによってひき起こされると考えられている種々の疾患における病原機構については、未だ十分には解明されていない。 H.pyloriのもつ病原因子のひとつに139kDaの空胞化毒素(VacA)があり、vacA遺伝子がコードしていることが知られている。空胞化毒素は、欧米では約50%の株が産生していると報告されており、また消化性潰瘍との関連が指摘されている。 患者血清中の抗VacA(空胞化毒素)抗体の測定系としては、血清の空胞化毒素に対する中和活性をみることにより評価した報告がいくつかみられる。日本においてもHiraiら20は胃癌患者6例の血清を調べて、全例に中和活性を認めたと報告している。しかし、中和活性をみる方法は、培養細胞の形態廟マ化として検出するため複雑であり、多数の検体を解析するには不適当である。 我々は、空胞化活性陽性株感染を示唆する血清中の抗VacA抗体をより簡便かつ正確に診断するため今回の研究を行なった。クローニングしたvacA遺伝子を大腸菌内で発現させ、組換えVacA蛋白を精製した。これを用いて、患者血清中の抗VacA抗体をイムノブロット法にて検出した。また、組換えVacA蛋白を家兎に免疫して得たrabbit抗VacA抗体を用いて菌からのVacA蛋白の検出および空胞化活性の測定も行なった。 [方法] 空胞化毒素産生株であるH.pylori株ATCC(American Type Culture Collection)43526からvacA遺伝子をクローニングした。上部消化管内視鏡検査を受けた68例から分離された株に対してVacAの蛋白検出を行なった。 H.pylori ATCC 43526よりDNAを精製し、vacA遺伝子ORF(open reading frame)全長(3864bp)をPCR(polymerase chain reaction)にて増幅した。PCR産物(3891bp)をプラスミドベクターにクローニングし、塩基配列を蛍光ダイデオキシ法にて決定した。さらに、制限酵素にて切り出したvacA遺伝子断片を融合蛋白発現用ベクターに組換えた(図1)。E.coliで発現させた組換えvacA蛋白を精製し以下の実験に用いた。 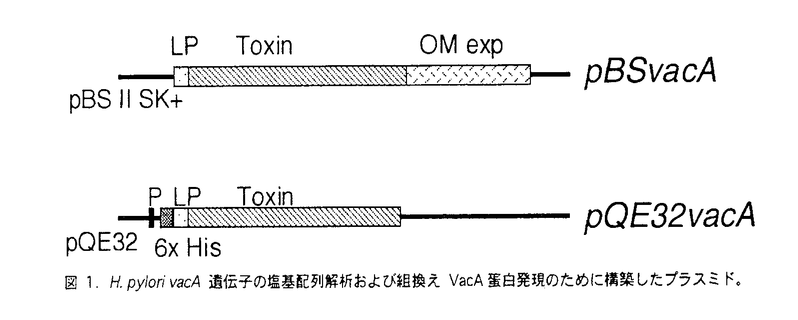 図1. H.pylori vacA遺伝子の塩基配列解析および組換えVacA蛋白発現のために構築したプラスミド。 図1. H.pylori vacA遺伝子の塩基配列解析および組換えVacA蛋白発現のために構築したプラスミド。 組換えVacA蛋白および合成ペプチドVA-1(アミノ酸配列:TSYKDSADRTTR、H.pylori ATCC 43526株VacAアミノ酸残基176-187)を家兎に免疫し、抗体を作製した。 作製した抗体を用いて、H.pylori培養上清または菌表面蛋白液よりVacA蛋白の検出を行った。SDS-PAGEにて検体を分離したのちnitrocellulose membraneに転写した。家兎抗血清と反応させたのち、125I-抗家兎IgG抗体を用いて検出した。 H.pylori培養上清を20倍に濃縮し、HeLa細胞培養液に添加した。24時間後に細胞の空胞化を光学顕微鏡で観察した。50%以上の細胞に空胞を認めた場合、空胞化毒素活性陽性と判定した。 上部消化管内視鏡検査を施行し、H.pylori感染診断(培養、迅速ウレアーゼ試験、胃液ureA-PCR法)がなされた97例より血清を採取した。血清中抗H.pylori IgG抗体価をELISA法により測定し、感染診断に併用した。 精製した組換えVacA蛋白を抗原としたイムノブロットを行い、患者血清中の抗VacA抗体を検出した。 [結果] 空胞化毒素陽性標準株であるATCC 43526のvacA遺伝子全長をクローニングし、全塩基配列を決定した。3873塩基のORFが存在し、1291アミノ酸をコードしていた。 H.pylori vacAの前半部分2233bpを制限酵素にて切り出し、蛋白発現用プラスミドに組換えた。E.coliで発現させ、精製した組換え蛋白はSDS-PAGEで予想される分子量をもつ単一のバンドとして認識された。 H.pylori株培養上清中のVacA蛋白の存在を組換えVacA蛋白および合成ペプチドVA-1に対して作製した家兎抗血清をもちいてイムノブロット法にて検討した。抗VA-1抗血清は空胞化毒素陽性株で抗組換えVacA抗血清と同じ大きさの蛋白を同定した。 患者血清中の抗VacA抗体の存在をイムノブロット法を用いて調べた。感染診断として、培養法と迅速ウレアーゼ試験、血清抗H.pylori抗体測定、および胃液PCR法を行い、2法以上において陽性であった場合にH.pylori感染陽性と判定した。97人中、84人はH.pylori感染陽性であった。血清中抗VacA抗体は97人中80人で陽性であり、H.pylori感染陽性患者では84人中77人(92%)、H.pylori感染陰性者では13人中3人(23%)において抗VacA抗体陽性であった(表1)。H.pylori感染と血清中抗VacA抗体の存在には有意な相関を認めた(p<0.0001;Fisher’s exact probability)。  表1 H.pylori感染と抗VacA抗体。 表1 H.pylori感染と抗VacA抗体。 H.pylori感染が確認された例で疾患別に検討すると、胃潰瘍、十二指腸潰瘍、慢性胃炎、内視鏡的健常者の各群において、抗VacA抗体陽性率はそれぞれ85%(22/26)、95%(20/21)、95%(18/19)、100%(14/14)であった(表2)。各群間で抗体陽性率には有意な差は認めなかった。 臨床分離H.pylori株68株中の59株(87%)にVacA蛋白を認めた。感染者の疾患別のVacA蛋白陽性率は、胃潰瘍、十二指腸潰瘍、慢性胃炎、胃癌でそれぞれ14/16(88%)、15/17(88%)、21/25(84%)、9/10(90%)であった。いづれの群でも陽性率は高値を示しており、各群間に統計的な有意差は認めていない。 空胞化毒素活性は68株中57株で陽性であった。分泌されたVacA蛋白の有無と比較すると、VacA蛋白陽性59株中54株(92%)、VacA蛋白陰性9株中3株(33%)で空胞化活性を認め、蛋白分泌の有無と活性の有無の間には関連が認められた(p<0.001;Fisher’s exact probability)。 [考察] H.pyloriの空胞化毒素はvacA遺伝子にコードされ、137kDaのvacA遺伝子産物は3領域にわけられる。すなわちN-末端の33アミノ酸からなるシグナルペプチド、約87kDaの空胞化毒素本体、およびC-末端の50kDaの領域である。組換えVacA蛋白は87kDaの空胞化毒素本体にほぼ一致するよう設計した。 過去の報告によると、空胞化毒素陽性株の感染が胃・十二指腸疾患の合併と関連があることが指摘されている。Figuraらは消化性潰瘍患者から分離した菌では24株中16株(67%)で空胞化活性を認めたのに対し、慢性胃炎の患者から分離した菌では53株中16株(30%)でのみ空胞化活性を認めたと報告している 空胞化毒素活性の測定は空胞化毒素の存在を解析するうえで、直接的な証明方法ではあるが、新鮮な胃生検組織からH.pyloriの培養を施行する必要があり、多数例での解析は困難である。患者血清中に存在する抗VacA抗体が空胞化毒素に対する中和活性をもつことに注目し、患者血清中の抗VacA抗体の有無を検討した報告もある。中和活性の測定は、複雑であり、結果に影響を与える要因が多い。それゆえ、我々は、本研究において、より簡便で正確な測定系の開発を試みた。 本研究において、我々はまずvacA遺伝子をクローニングし、E.coli内で組換えVacA蛋白を大量に発現させた。組換えVacA蛋白を用いて作製した抗体は、H.pyloriが分泌する空胞化毒素を特異的に認識した。 精製した組換えVacA蛋白を用いたイムノブロット法により、H.pylori感染者の血清中には高頻度に抗VacA抗体が存在することが明らかになった。さらに、感染者の疾患に関わらずいづれの群でも高い抗体陽性率を認めた。この結果から、日本においては欧米と異なり、空胞化毒素産生株が大多数を占めることが示唆された。 さらに、空胞化毒素産生株の感染頻度を確認するため、我々は菌表面蛋白中のVacA蛋白の同定および空胞化毒素活性の確認を行なった。その結果、68株中59株(87%)の株はVacA蛋白を分泌しており、また、57株(84%)の培養上清は空胞化活性を示した。データからも日本においては空胞化毒素産生株が高率に感染していることが示唆された。 胃癌は現在も世界中でも高頻度にみられる悪性疾患のひとつであるが、日本は特に胃癌の発生が多い国である。疫学的研究により胃癌患者では、H.pylori感染が高率に認められており、近年動物実験でも、H.pylori感染動物で胃癌が発生することが報告されている。我々の結果は、日本人のH.pylori感染者の大半は空胞化毒素陽性株に感染していることを示しているが、日本で胃癌の発生が多い原因は、H.pyloriの感染率が高いことに加えて、空胞化毒素産生株が多いことが関連している可能性があるものと考えられる。今回、抗体測定系を作製したが、こうした系を用いて大規模な前向き研究を行なうことにより、空胞化毒素産生H.pylori感染と胃癌の発生について解明できる可能性があるものと思われる。 |