血小板由来増殖因子(platelet-derived growth factor:PDGF)は線維芽細胞、平滑筋細胞、グリア細胞などの間葉系細胞に対して増殖活性を持つ因子として、ヒト血小板より精製された。その後、PDGFは、血小板のみならず、血管内皮細胞、マクロファージ、骨芽細胞など多くの細胞より分泌、産生されることが明らかとなり、さらに、細胞遊走活性や細胞外マトリックス産生の刺激など多彩な生理作用が報告されている。さらに、間葉系細胞以外にも、血管内皮細胞や神経細胞など多種の細胞種がPDGFの標的になることが知られている。また、PDGF B鎖はサル肉腫ウイルスの癌遺伝子v-sisの産物と同一であることが判明し増殖因子産生と密接に関連した発癌機構の存在を示唆するものとして注目された。近年では創傷治癒や動脈硬化病変の形成にもPDGFが重要な役割を果たすと考えられている。 このように、PDGFはさまざまの細胞に対する多様な生理作用を持つことが明らかにされつつあり、その細胞内分子機構の解明にも精力が注がれているが、心筋細胞に対する作用に関する報告は少ない。近年、PDGF B鎖ノックアウトマウスにおいて、胎生期に心・大血管の形態異常を認めたことから、PDGFが胎生期の心臓の発達に何らかの役割を果たしていることが示唆された。そこで、私はニワトリ胚(10日)培養心筋細胞を用い、PDGFが心筋細胞に対しても直接作用し、増殖を促進するかどうか、そしてPDGFが心筋細胞に対して作用するのであれば、その細胞内分子機構がいかなるものであるかを解析した。 まずPDGFの心筋細胞に対する増殖効果を解析するため、MTT[3-(4,5-dimethylthiozol-2-yl)-2,5-diphenyltetrazolium bromide]法を用いて生細胞数を、BrdU(5-bromo-2’-deoxyuridine)の取り込み量でDNA合成能を評価した。PDGFはA鎖、B鎖の2種類のポリペプチド鎖が二量体化した構造をとり、PDGF-AA,-AB,-BBの3種類のアイソフォームが存在するため、それぞれについて増殖効果を検討した。PDGF-ABおよび-BBは用量依存性に生細胞数を増加させたが、PDGF-AA投与では有意な変化は認められなかった(5ng/ml PDGF-AA,-AB,-BB:101±4%,115±4%,122*±4%,n=4,*P<0.05)。DNA合成能に関してもMTT法の結果と同様、PDGF-ABおよび-BBの投与で合成能が亢進したが、PDGF-AAによる変化は認めなかった(5ng/ml PDGF-AA,-AB,-BB:104±11%,202*±18%,295*±25%,n=4,*P<0.05)。この結果からPDGFの心筋細胞に対する増殖作用にはPDGF B鎖が重要であることが示唆された。また、PDGFの受容体には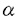 サブユニットと サブユニットと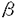 サブユニットがあり、B鎖は,両サブユニットと結合するが、A鎖はサブユニットとのみと結合する。そこでリガンド結合法(Scatchard解析)を用いそれぞれのアイソフォームの結合しうる受容体総数(Bmax)を解析したところ、PDGF-AA:5±1,-AB:63±12,-BB:126±24(fmol/106cells)となり、各アイソフォームの作用の相違は、結合しうる受容体数に依存しており、PDGF-AAに増殖作用を認めないのは結合する受容体数が極めて少ないためと推察された。 サブユニットがあり、B鎖は,両サブユニットと結合するが、A鎖はサブユニットとのみと結合する。そこでリガンド結合法(Scatchard解析)を用いそれぞれのアイソフォームの結合しうる受容体総数(Bmax)を解析したところ、PDGF-AA:5±1,-AB:63±12,-BB:126±24(fmol/106cells)となり、各アイソフォームの作用の相違は、結合しうる受容体数に依存しており、PDGF-AAに増殖作用を認めないのは結合する受容体数が極めて少ないためと推察された。 以上の結果より、PDGFは心筋細胞にも作用し増殖を促進することが示されたが、さらに、その細胞内分子機構に関しPDGF-BBを用いて解析した。まず、細胞増殖・分化に関与すると考えられる初期応答遺伝子c-fos mRNAの発現をノーザンブロット法を用いて解析したところ、PDGF-BB投与後30分にc-fos mRNAの一過性の発現を認めた。また、c-Fosやc-Junなどの複合体であり、さまざまな遺伝子の発現に関わる転写因子AP(activator protein)-1の結合活性能をゲルシフト法により検討したところ、PDGF-BBにより有意にその結合活性能が上昇した。このc-fos mRNAの発現およびAP-1結合活性能上昇はチロシンキナーゼ阻害薬(100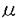 M genistein)およびMAPK(mitogen-activated protein kinase)kinase阻害薬(10or50M PD98059)により有意に抑制された。一方、PKC(protein kinase C)阻害薬(250nM calphostin C)はPDGF-BBによるAP-1結合活性能の上昇をある程度抑制したもののc-fos mRNAの発現には全く影響しなかった。MAPK kinase阻害薬がc-fos mRNAの発現およびAP-1結合活性能上昇を抑制したことからPDGF-BBのシグナリングにMAPKの活性化が重要であると考えられたため、実際にウエスタンブロット法および免疫染色にてMAPKの活性化について解析したところ、PDGF-BB投与後直ちに、MAPKのリン酸化および核内への移行が起こることが示された。 M genistein)およびMAPK(mitogen-activated protein kinase)kinase阻害薬(10or50M PD98059)により有意に抑制された。一方、PKC(protein kinase C)阻害薬(250nM calphostin C)はPDGF-BBによるAP-1結合活性能の上昇をある程度抑制したもののc-fos mRNAの発現には全く影響しなかった。MAPK kinase阻害薬がc-fos mRNAの発現およびAP-1結合活性能上昇を抑制したことからPDGF-BBのシグナリングにMAPKの活性化が重要であると考えられたため、実際にウエスタンブロット法および免疫染色にてMAPKの活性化について解析したところ、PDGF-BB投与後直ちに、MAPKのリン酸化および核内への移行が起こることが示された。 近年、PDGFを含む増殖因子やサイトカインの細胞内情報伝達物質として転写因子STATs(the activation of signal transducers and activators of transcription)が重要であることが示されており、PDGFの心筋細胞に対する作用にも寄与している可能性が考えられた。しかしながら、免疫沈降およびウエスタンブロット法による解析の結果、STAT3はPDGF投与前から既に活性化されており、PDGF投与後も変化を認めなかった。STAT1に関しては、PDGF投与前後通してその活性化は認めなかった。このような他の細胞とのSTATs活性化状態の相違は、種、組織、成長段階の相違に起因するものと推察された。また、PDGFの細胞内分子機構にはPLC(phospholipase C)-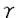 を介した細胞内カルシウム動員も報告されているが、ニワトリ胚培養心筋細胞においては、indo-1を用いて測定した細胞内カルシウム濃度もパッチクランプ法を用いて測定したLタイプカルシウムチャンネル流入量もPDGF投与前後で不変であった。 を介した細胞内カルシウム動員も報告されているが、ニワトリ胚培養心筋細胞においては、indo-1を用いて測定した細胞内カルシウム濃度もパッチクランプ法を用いて測定したLタイプカルシウムチャンネル流入量もPDGF投与前後で不変であった。 このように、細胞内分子機構に関する解析の結果、MAPK cascadeがPDGFの心筋細胞に対する作用発現に重要な役割を果たしていることが示唆された。そこで、最後に前記の3種のキナーゼ阻害薬の増殖作用への影響をBrdUの取り込みで評価した。阻害薬無投与ではPDGF-BBは有意にDNA合成能を促進した(292*±19%,n=6,*P<0.05vs.control)。PKC阻害薬(250nM Calphostin C)はPDGF-BBによるDNA合成促進に有意な影響は与えなかった(264±33%)。MAPK kinase阻害薬(PD98059)は用量依存性にDNA合成促進を抑制した(10M:198*±14%,50M:168*±14%,n=6,*P<0.05vs.PDGF-BB-treated cells)。PDGF-BBによるDNA合成促進はチロシンキナーゼ阻害薬(100M genistein)により完全に抑制された(85*±5%,n=6,*P<0.05vs.PDGF-BB-treated cells)。このことから、PDGFによるチロシンキナーゼおよびMAPKの活性化が心筋細胞の増殖を導くことが示された。 以上、結論として、PDGF(-AB,-BB)はニワトリ胚培養心筋細胞に直接作用し、増殖効果を発現した。その細胞内分子機構においてはMAPKの活性化を介したc-fos mRNAの発現や転写因子AP-1の結合活性能亢進が重要であることが示された。 |