造血幹細胞から各種血球への分化・増殖には様々なサイトカインが関与しており、その生理活性と標的細胞により大きく(1)lineage-nonspecific early-acting factorと(2)lineage-specific late-acting factorに分類される。造血幹細胞の分化・増殖は単独のサイトカインでは支持されず、両者の組み合わせが必要であることが示されている。赤芽球系細胞への分化・増殖には(1)のうちのstem cell factor(SCF)と(2)のうちのerythropoietin(EPO)の組み合わせが最も効果的であることが知られている。しかし両者の組み合わせの必要性、両者の組み合わせによる相乗効果の発現の仕組み等は不明である。造血幹細胞/前駆細胞はその分化に伴い、(1)に対する受容体がdown-regulateされ、その感受性を失い、かつ(2)に対する受容体が発現され、その感受性を獲得することが一部のサイトカインについて示されており、両者の受容体が共に発現している前駆細胞において相乗効果が現れるものと思われる。 ヒト骨髄細胞をEPOを加えた半固形培地で培養すると経過に伴い2種類の赤芽球コロニーが出現する。即ち培養5〜7日目の小さなコロニーと10日目以降の花火がはじけたようなバーストである。前者を形成する元の細胞をCFU-E、また後者の元の細胞をBFU-Eと称し区別される。BFU-Eは造血幹細胞から赤芽球系細胞へと分化した前駆細胞であり、CFU-EはBFU-Eよりもさらに成熟した前赤芽球に近い段階にある前駆細胞である。BFU-Eレベルの前駆細胞は、いまだEPO受容体(EPOR)の発現が少なくEPOに対する感受性を有しないが、分化に伴いその発現を増し、CFU-Eレベルの前駆細胞でEPOに対する感受性が最大になると報告されている。赤芽球系細胞の分化・増殖におけるSCFの重要性は、SCF又はその受容体であるc-kitををコードする遺伝子に変異を有するS1/S1d、W/Wvマウスが共に重症の巨赤芽球性貧血を呈することから示されている。その原因として、CFU-Eレベルの赤芽球系前駆細胞が減少していることが報告された。 IL-6受容体はIL-6に固有な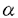 鎖(IL-6R)とIL-11等と共通の 鎖(IL-6R)とIL-11等と共通の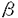 鎖(gp130)とから構成されていることが知られている。最近の研究で、臍帯血から精製したCD34+細胞の大部分がgp130を発現しているのに対して、IL-6Rは30-50%にしか発現していないことが報告された。可溶性IL-6受容体鎖(sIL-6R)とIL-6の組み合わせを用いると、IL-6Rを発現していない細胞に対してもgp130を活性化し、IL-6と同等の細胞内シグナル伝達を引き起こすことが知られている。最近sIL-6RとIL-6の組み合わせを用いたgp130の活性化と、SCFによるc-kit活性化により、EPO非存在下にCD34+細胞より成熟赤血球が産生されることが見出された。この結果は、造血幹細胞から赤芽球系細胞への分化にはEPOの受容体であるEPORは不必要で、赤芽球系前駆細胞上に発現しているEPOR以外のサイトカイン受容体を介する刺激により、その増殖と生存を保つことで起こることを示唆する。一方、c-kitとEPORを共発現する細胞株で、SCF刺激によりc-kitがEPORをリン酸化することが報告され、赤芽球系細胞の分化・増殖におけるSCFとEPOの協調作用の仕組みの一つとしての仮説が提唱された。このモデルが当てはまるとすると、c-kit、gp130同時刺激によるEPO非存在下の赤血球造血においても、c-kitからEPORを通じて分化のシグナルが伝達される可能性は否定できない。EPO存在下における赤血球造血においても、sIL-6R・IL-6を用いたEPO非存在下における赤血球造血においても、SCF/c-kit系は大変重要な役割を担っていると考えられる。 鎖(gp130)とから構成されていることが知られている。最近の研究で、臍帯血から精製したCD34+細胞の大部分がgp130を発現しているのに対して、IL-6Rは30-50%にしか発現していないことが報告された。可溶性IL-6受容体鎖(sIL-6R)とIL-6の組み合わせを用いると、IL-6Rを発現していない細胞に対してもgp130を活性化し、IL-6と同等の細胞内シグナル伝達を引き起こすことが知られている。最近sIL-6RとIL-6の組み合わせを用いたgp130の活性化と、SCFによるc-kit活性化により、EPO非存在下にCD34+細胞より成熟赤血球が産生されることが見出された。この結果は、造血幹細胞から赤芽球系細胞への分化にはEPOの受容体であるEPORは不必要で、赤芽球系前駆細胞上に発現しているEPOR以外のサイトカイン受容体を介する刺激により、その増殖と生存を保つことで起こることを示唆する。一方、c-kitとEPORを共発現する細胞株で、SCF刺激によりc-kitがEPORをリン酸化することが報告され、赤芽球系細胞の分化・増殖におけるSCFとEPOの協調作用の仕組みの一つとしての仮説が提唱された。このモデルが当てはまるとすると、c-kit、gp130同時刺激によるEPO非存在下の赤血球造血においても、c-kitからEPORを通じて分化のシグナルが伝達される可能性は否定できない。EPO存在下における赤血球造血においても、sIL-6R・IL-6を用いたEPO非存在下における赤血球造血においても、SCF/c-kit系は大変重要な役割を担っていると考えられる。 ヒト赤芽球分化におけるこのc-kitの役割を解析するのが本実験の目的である。 研究方法 SCFによるEPOR遺伝子発現誘導の解析については、SCF存在下にはじめてEPO感受性を獲得するヒト白血病細胞株HML/SEを使用し、各種サイトカイン存在下における分化・増殖をMTT assay、免疫染色、フローサイトメーター等を用いて検討した。またSCF存在下におけるEPOR mRNAの発現をNorthern blotting法で解析した。更にレポーター遺伝子一過性発現系でSCF存在下におけるEPORプロモーターの活性化、及び活性化に寄与する転写因子結合配列を評価した。 c-kit、gp130同時刺激によるEPO非存在下の赤血球造血のメカニズムの解析においてはヒト臍帯血より磁気ビーズ法にてCD34+細胞を精製し、使用した。EPORに対するアンチセンスオリゴDNAを培養液中に添加し、EPORの発現を抑制することでc-kit、gp130同時刺激による赤血球造血におけるc-kitとEPORの相互作用及び役割を解析した。赤芽球分化の評価はdianisidineによるヘモグロビン染色によった。また培養により発現する遺伝子をRT-PCR法で検討した。 結果及び考察 (1)ヒト白血病細胞株HML/SEにおけるSCF刺激によるエリスロポエチン受容体遺伝子の発現誘導と、エリスロポエチンに対する感受性の獲得 ヒト白血病細胞株HML/SEの増殖を単独で支持できるサイトカインはgranulocyte-macrophage colony-stimulating factor(GM-CSF)とSCFであり、EPOはいかなる濃度においても増殖に対する効果は認められなかった。しかし、SCFの共存下でEPOは用量依存的にHML/SE細胞の増殖を刺激した。またGlycophorin A、ヘモグロビンの産生など、赤芽球系への分化を誘導した。このEPOの作用はGM-CSFとの組み合わせでは認められなかった。SCFがEPORを発現していないHML/SE細胞に対して、EPORの発現を誘導しEPOに対する感受性を獲得させたことが考えられたので、Northern blotting法でSCF存在下におけるHML/SE細胞のEPOR mRNAの発現を調べた。その結果、SCF刺激により経時的にEPOR mRNAの発現が増強することが認められた。EPOR遺伝子の発現には、そのプロモーター中のGATA及びSp1結合配列が重要であることが報告されているため、レポーター遺伝子一過性発現系を用いてSCF刺激によるEPORプロモーターの活性化を評価した。その結果、SCF刺激によってEPORプロモーターは強く活性化されることが明らかとなった。GATA及びSp1結合配列に変異を導入し、SCF刺激によるEPORプロモーターの活性化におけるそれぞれの転写因子の寄与を評価した。その結果SCF刺激ではGATA結合配列よりもSp1結合配列が重要であることを示唆する結果を得た。このことは、SCFがGATA-1によって転写が制御されている他の赤芽球系特異的遺伝子の発現を誘導せずに、EPOR遺伝子の発現のみを特異的に誘導する理由と考えられた。 (2)ヒト臍帯血由来CD34陽性細胞からの、c-kit、gp130同時刺激によるEPO非存在下における赤血球造血のメカニズム ヒト臍帯血CD34+細胞をSCF、sIL-6R、IL-6の共存下に培養すると赤芽球が生じるが、この時発現している遺伝子をRT-PCR法で検討すると、EPO存在下に生じた正常赤芽球と同様に、EPORを含め赤芽球系特異的遺伝子の発現が認められた。EPOR mRNAに対するアンチセンスオリゴDNAを培養に加えEPORの発現を抑制すると、EPO依存性の赤血球造血のみならず、c-kit、gp130同時活性化によるEPO非存在下の赤血球造血も阻害された。この原因として前述のc-kitによるEPORのリン酸化の他に、SCF、sIL-6R、IL-6存在下で培養したCD34+細胞がEPOを産生することが考えられた。この鑑別のため、RT-PCR法にてSCF、sIL-6R、IL-6存在下で培養したCD34+細胞中のEPO mRNAの発現を調べた。その結果、特に赤芽球系細胞においてEPO mRNAが発現していることを示唆する結果を得た。高濃度の抗ヒトEPO中和抗体をSCF、sIL-6R、IL-6存在下で培養しているCD34+細胞の培養液中に添加することにより、赤芽球分化は完全に阻害された。以上の結果より、SCF、sIL-6R、IL-6存在下において赤芽球系前駆細胞はEPOを自己分泌することにより、分化・成熟することが示唆された。また、c-kit、gp130同時活性化によるEPO非存在下の赤血球造血においてc-kitがEPORをリン酸化することにより赤芽球分化シグナルを伝達する可能性は否定された。 結論 (1)SCFがCFU-E以前の段階の幼若な前駆細胞に対してEPORの発現を誘導しEPOに対する感受性を獲得させることを示唆する結果を得た。これはSCFとEPOが協調的に作用するメカニズムの一つと考えられた。 (2)EPORプロモーターの解析より、SCFによるEPOR遺伝子発現誘導においては、GATA結合配列よりもSp1結合配列が重要であることを示唆する結果を得た。 (3)ヒト臍帯血CD34+細胞からのc-kit、gp130同時刺激によるEPO非存在下の赤血球造血は、赤芽球系前駆細胞におけるEPOの自己分泌によることが示唆された。 |