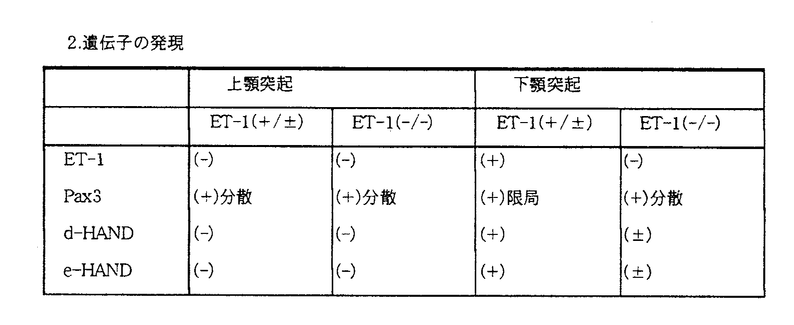
本研究はヒトもしくはマウスなどの哺乳類の下顎発生メカニズムを明らかにするため、顎顔面奇形モデルであるエンドセリン-1遺伝子欠損マウスを用いて、下顎ならびに口腔発生過程を詳細に観察するとともに、顎顔面を構成する主要成分となる神経堤細胞の分化過程と、それを制御する遺伝子発現との関連についての解析を試みたものであり、下記の結果を得ている。 1.肉眼像:ET-1(-/-)とET-1(+/+,+/-)についてその表現型を詳細に比較、検討したところ、ET-1(-/-)ではすでに報告のある全例下顎正中癒合不全、小顎化、耳介の低形成の他、下顎にET-1(+/+,+/-)では存在しない3列の洞毛(触覚毛、ヒゲ)の毛根(whisker pad)が新たに認められた。この形態は、上顎突起由来の上顎の3列の洞毛形態と類似していた。 2.神経像:神経の免疫染色からはET-1(-/-)では異所性(尾側)に過剰な(複数の)下顎神経束が下顎異所性に生じた洞毛のwhisker pad様構造物へと伸びているのが観察され、この下顎神経の異常形態は、上顎神経の鼻尖末端部の神経形態とよく類似していた。 3.骨・軟骨像:胎生15.5日以降の骨・軟骨標本からは、メッケル軟骨は腹側先端には認められたが軸部では完全に欠失し、下顎骨の形態が頬骨形態と類似していることが明らかになった。 4.血管像:バリウム注入による血管描出からは、ET-1(-/-)では、顔面動脈から分枝した下唇動脈過剰枝が4本外内側に枝分かれして表層に広く分布し、下顎に存在するwhisker pad様構造物へと走行しており、この複数の枝分かれのために、上唇動脈へと走行する枝はET-1(+/+,+/-)と比較するとかなり細くなっていたが、上唇動脈は、顎動脈からの分枝と吻合することによって再び太くなり、血液供給を補っているものと示唆された。 5.組織像:トルイジンブルー染色による組織切片像からは、胎生10.5日ET-1(+/+,+/-)の頭側咽頭弓上皮直下の間葉細胞に球状に肥大した核、細胞間基質の増大、多数の間葉細胞の凝集が認められたが、ET-1(-/-)では認められず、その疎な細胞形態は上顎突起の組織像と類似していた。また、胎生16.5日ET-1(-/-)ではメッケル軟骨軸部、腺組織が欠失し、過剰血管、神経が認められ、これらは前述した1〜4の結果と一致していた。 6.遺伝子の発現:胎生初期胚のWhole mount in situhybridizationにより、ET-1(+/+,+/-)の下顎突起では、上顎突起ではPax3優位、下顎突起ではHAND優位であったのに対し、ET-1(-/-)の下顎突起ではHANDの発現が著明に減少し、相対的にPax3の発現領域が拡大していることが明らかになった。 以上、本論文はET-1(-/-)の形態異常が下顎突起由来の構造物が上顎突起によく類似した形態をとることから、下顎の形態の独自性を与えている可能性があるというET-1の新たな役割について、およびそのメカニズムとしてET-1が、上皮-間葉相互作用のメディエーターとして咽頭弓の上皮直下に遊走してきた神経堤細胞に由来する外胚葉性の間葉細胞に作用し、dHAND,eHAND,Pax3の発現を調節することによって下顎の形態の発達を促進しているという新しい知見を明らかにした。本研究はこれまで未知に等しかった胎生初期の顎発生におけるメカニズムの解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |