本論文は3章からなり、第1章は、ニンジン直接不定胚形成を利用した迅速かつ簡便な遺伝子導入系を開発について、第2章は、この系を用いてのCHB1遺伝子の胚形成における働きの解析について、第3章は、第1章で開発した遺伝子導入系を用いてのCHB2遺伝子の胚形成における働きの解析について述べられている。 近年、シロイヌナズナの突然変異体をもとに、胚のパターン形成に関係する遺伝子が単離され、その解析から胚形成の分子機構の一部が明らかになりつつある。しかし、この方法を用いた胚形成機構の研究には、多くの変異体が胚致死でエッセンシャルな遺伝子の単離が必ずしも容易でないことや、大量の接合子胚の操作を必要とする生理、生化学的解析の難しさ等の問題点もある。このような問題を解決し、胚形成機構を多面的に解析するためには、異なるアプローチが求められる。こうした中で、ニンジン不定胚形成はin vitroで接合子胚とほぼ同じ過程で進行し、同調的かつ高頻度に胚分化が起こることから、胚形成のもうひとつのすぐれたモデル系と考えられている。この実験系を用いて多くの生理、生化学、分子生物学的な解析がなされ、不定胚形成特異的な遺伝子もいくつか単離されてきている。しかし、効率的な遺伝子導入系がないために、遺伝子の機能解析にまで到っている例は少なかった。 そこで、第1章として、論文提出者は、論文提出者自身が修士課程で開発したニンジン直接不定胚形成系を利用することで、迅速かつ簡便なニンジン不定胚への遺伝子導入系の確立を試みた。マーカーとしては35Sプロモーターの下流に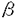 -グルクロニダーゼ(GUS)遺伝子をつないだ融合遺伝子とハイグロマイシン耐性遺伝子を、遺伝子導入法としてはアグロバクテリアを介した方法を、植物材料としてはてニンジン胚軸外植片を用いて、様々な条件を検討した。その結果、効率よく遺伝子導入不定胚を作成する手法の開発に成功した。この成功はこれまで分子機構の解明が困難であったニンジン不定胚形成に逆遺伝学的な方法の導入を可能にし、その研究の飛躍的な進展ももたらす契機を与えた。 -グルクロニダーゼ(GUS)遺伝子をつないだ融合遺伝子とハイグロマイシン耐性遺伝子を、遺伝子導入法としてはアグロバクテリアを介した方法を、植物材料としてはてニンジン胚軸外植片を用いて、様々な条件を検討した。その結果、効率よく遺伝子導入不定胚を作成する手法の開発に成功した。この成功はこれまで分子機構の解明が困難であったニンジン不定胚形成に逆遺伝学的な方法の導入を可能にし、その研究の飛躍的な進展ももたらす契機を与えた。 第2章では、論文提出者は、第1章で確立した遺伝子導入系を用いて、胚形成過程で強く発現する転写制御因子CHB1(HD-Zip1タイプ)の解析を行った。35Sプロモーターの下流にCHB1 cDNAをセンスおよびアンチセンス方向につなぎ、ニンジン不定胚に導入した。遺伝子導入した不定胚の形態を調べたところ、センス鎖を導入したものでは80%で子葉が融合して大きくなるか、あるいは子葉状の構造が多数形成されるという変異が見られた。一方、アンチセンス鎖を導入したものでは子葉と茎頂を含む胚の上部器官の発達が抑制されることがわかった。以上の結果と胚形成初期にCHB1 mRNAは胚上部に局在するという観察から、論文提出者はCHB1は不定胚形成において子葉と茎頂分裂組織を含む胚の上部器官の形成を制御しているという仮説を提出した。また、CHB1をアンチセンス方向に導入した形質転換幼植物体から二次不定胚を誘導したところ、不定胚形成が阻害され、カルスが形成された。この結果は、CHB1が胚形成過程のみならず、不定胚誘導にも働いていることを示すものであった。これらの結果は、HD-Zip1ホメオボックス遺伝子が胚形成の進行と誘導に関与していることを初めて示したもので、高く評価される。 さらに第3章では、論文提出者は、心臓型胚で一過的に発現が上昇するホメオボックス遺伝子CHB2(HD-Zip1タイプ)の発現と機能の解析を行った。in situでの発現を調べたところ、心臓型胚では子葉原基に発現し、魚雷型胚以降は維管束が形成される領域に局在することがわかった。次に、CHB2をアンチセンス方向に導入した胚を作成したところ、形質転換胚では球状胚から心臓型胚への発達が特異的に阻害されることが明らかになった。この結果よりCHB2は初期心臓型胚の子葉原基の形成を制御していることが示唆された。これらのデータは、子葉原基の形成を探る上での重要な知見となった。 ここに得られた結果の多くは新知見であり、いずれもこの分野の研究の進展に重要な示唆を与えるものであり、かつ本人が自立して研究活動を行うのに十分な高度の研究能力と学識を有することを示すものである。よって、得字圭彦提出の論文は博士(理学)の学位論文として合格と認める。 | 

 m。(G)-(M) Bar=100
m。(G)-(M) Bar=100