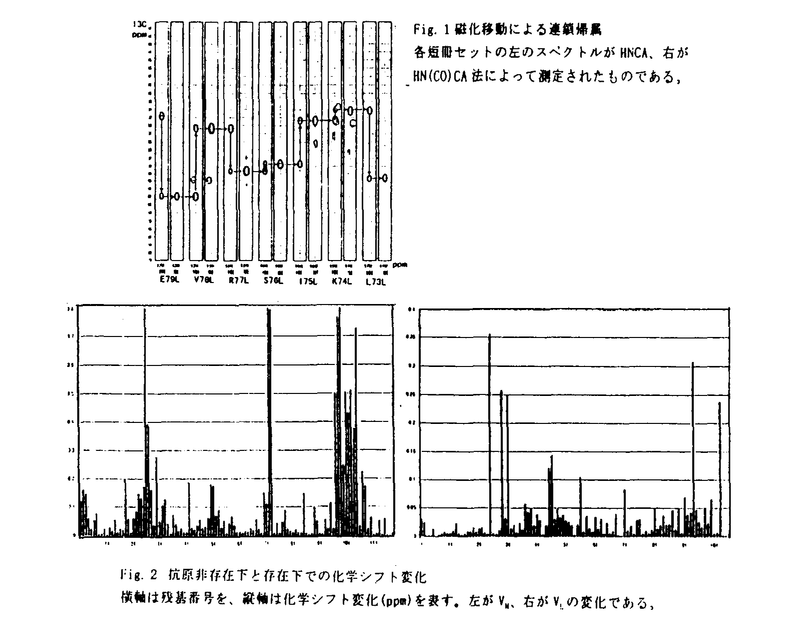
免疫グロブリン、B細胞、T細胞の抗原レセプターおよびサイトカインレセプターなど、免疫グロブリンスーパーファミリーに属する蛋白質は、逆平行 シートが2層に対向したイムノグロブリンフォールド(Igフォールド)と呼ばれる立体構造上の基本構造から構成されている。免疫応答、サイトカインの認識、シグナル情報の伝達など生命現象にとって重要な機能はこのIgフォールドを土台にして発現される。一方、Igフォールドを利用した抗癌剤、例えばFv fusion蛋白質やimmunotoxinsの開発も行われ、臨床応用の観点から、水溶液中におけるIgフォールドのより高い安定性が望まれている。 シートが2層に対向したイムノグロブリンフォールド(Igフォールド)と呼ばれる立体構造上の基本構造から構成されている。免疫応答、サイトカインの認識、シグナル情報の伝達など生命現象にとって重要な機能はこのIgフォールドを土台にして発現される。一方、Igフォールドを利用した抗癌剤、例えばFv fusion蛋白質やimmunotoxinsの開発も行われ、臨床応用の観点から、水溶液中におけるIgフォールドのより高い安定性が望まれている。 本研究では、ハイブリドーマ由来の抗ダンシルリジン抗体を酵素消化して得られるFvフラグメント(VH、VLヘテロダイマー)を研究対象として取り上げ、動物細胞を用いたFvフラグメント発現系における均一安定同位体標識法およびそのNMRシグナル帰属法の確立し、生物学的および抗体工学的にも重要なIgフォールドの動的立体構造の解析を行っている。 序論に続く第2章では、動物細胞を用いたFvフラグメント発現系における均一安定同位体標識法が述べられている。分子量25KのFvフラグメントの立体構造をNMRを用いて解析するためには、安定同位体原子をFvフラグメントに導入することが必須である。動物細胞による発現系では、通常、原核細胞で用いられている最小培地による均一安定同位体標識法が適応できない。そこで、15N、13C標識クロレラ藻体由来の蛋白質酸加水分解物を培地に加え細胞培養することにより、均一安定同位体標識を行った。しかしながら、酸加水分解物に含まれる細胞毒性成分のため細胞が死滅し、充分な量の蛋白質を調製できなかった。酸加水分解物の精製法を詳細に検討した結果、細胞毒性成分の効果的除去法の確立に成功した。また、用いる培地にはGlnが0.5g/lと大量に必要とされるため、安定同位体標識Glnの調製法に着手した。Glu生産菌の野生株を種々の条件で培養を行い、必要十分量のGlu生産に適した条件を決定した。さらに培地からMn2+を除去することにより醗酵生産物がGluからGlnへ平均80%の効率で転換することを発見した。得られた安定同位体標識Glnを精製された蛋白質酸加水分解物に加え、細胞培養を行い、均一安定同位体標識Fvフラグメントの大量調製に成功した。 第3章では、均一安定同位体標識Fvフラグメントの主鎖1H-15N HSQCシグナルを、アミノ酸タイプ別および部位特異的に帰属している。従来、動物細胞を用いた安定同位体蛋白質のNMR研究では、代謝により互いに変換するアミノ酸(GlyとSer残基など)のアミノ酸タイプ別帰属は不可能であると考えられてきた。学位申請者は、GlyおよびSer残基のC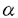 化学シフト値が大きく異なることに着目し、HNCA測定条件を検討することにより、GlyとSer残基シグナルの分離観測に成功した。さらに、GlnとGlu残基、AsnとAsp残基由来のシグナルに対してもアミノ酸タイプ別帰属に成功した。 化学シフト値が大きく異なることに着目し、HNCA測定条件を検討することにより、GlyとSer残基シグナルの分離観測に成功した。さらに、GlnとGlu残基、AsnとAsp残基由来のシグナルに対してもアミノ酸タイプ別帰属に成功した。 以上のアミノ酸タイプ別帰属に基づき、NOEならびに磁化移動に基づく連鎖帰属法を行い、ハプテン存在下および非存在下の主鎖シグナルの部位特異的帰属も確立した。 第4章では、ペプチド主鎖に対する緩和時間測定と重水素交換実験の結果をもとに、Igフォールド(VH、VLドメイン)の水溶液中における動的立体構造を議論している。 15N緩和時間(T1、T2)および1H-15N間のNOEの測定から、1H-15Nベクトルの空間的な制約性を示すオーダーパラメータを算出したところ、両ドメインに対し、ナノ秒からマイクロ秒の運動性は同じであることが判明した。さらにハプテンの有無も両ドメインの運動性に影響を与えないことを明らかにした。 一方、ハプテン非存在下の主鎖アミド水素重水素交換実験より、VHドメインはVLドメインに比べてより柔軟性が乏しいことが判明した。アミノ酸のhydropathic indexに基づき両ドメインの疎水性解析を行ったところ、界面を形成するVL側領域は多くの親水性残残基から構成されているのに対し、VH側領域は疎水性アミノ酸が多く存在していることが示された。したがって、この親水性・疎水性アミノ酸の分布に違いが、両ドメインの柔軟性に関与していると考えた。次に、ハプテン存在下で同様な測定を行ったところ、両ドメインとも界面形成領域残基の交換速度定数が1〜2桁以上低下することが示された。ハプテン添加に伴い界面形成領域に化学シフト変化が誘起されることと考え合わせ、ハプテン結合によりVH-VLドメイン間の相互作用が変化し、その結果、両ドメインの会合状態が安定化されると考えた。 以上、本研究は動物細胞を用いた蛋白質発現系における均一安定同位体標識法を確立し、高分子量蛋白質の主鎖NMRシグナル帰属の戦略を示し、さらにIgフォールドの動的立体構造を解析したものである。本研究で確立した手法および得られた知見は、動物細胞による蛋白質発現系を用いた構造生物学的研究やIgフォールドを用いたタンパク工学的研究の基礎に貢献し、博士(薬学)の学位に値すると判定した。 |