本研究は、血管内皮細胞におけるシェアストレス応答性転写因子群(AP-1、CREB-1及びCREM-1、NF-kappaB)を介した遺伝子発現誘導に関与する細胞内情報伝達機構を解析し、流れ負荷による遺伝子発現誘導のシェアストレス依存性、チロシンキナーゼのシェアストレス応答性転写因子群を介した転写誘導への関与、さらにFAK(Focal Adhesion Kinase)のシェアストレス依存的遺伝子発現誘導への関与を検討したものであり、以下の結果を得ている。 1)シェアストレス応答配列(TRE、CRE、kappaB)のルシフェラーゼ・リポーターを培養ウシ胎仔大動脈内皮細胞(BFAE)にトランスフェクションし、平行平板型流れ負荷装置を用いて動脈レベルのシェアストレス(15dyne/cm2)が加わるように流れ負荷した際の転写活性の変化を検討した。その結果、TRE、CRE、kappaB共に負荷後3時間から有意に発現上昇が認められ、12時間後にピークに達した。負荷後24時間にはbasal levelを若干上回る程度まで減少した。 2)流れ負荷を行った際、細胞に加わる刺激はシェアストレスだけでなく、液性因子の到達量の増加もある。この二つの効果を分離して検討する為、シェアストレスは粘度に比例し、拡散定数は粘度に反比例することに着目し、流速一定の条件で灌流液の粘性を変化させて流れ負荷によるTRE転写誘導がシェアストレス依存性か液性因子依存性であるかを検討した。その結果、低粘度、高粘度いずれも流速依存的な転写誘導が認められたが、流速一定で常に高粘度の方が低粘度よりも高い転写誘導を示した。さらに、各条件下でのシェアストレスを計算してプロットしたところ、一本の曲線となり流れ負荷による転写誘導がシェアストレス依存性であることが示された。 3)シェアストレス依存的なTRE、CRE、kappaBを介した転写誘導に関与する転写制御因子を特定する為、静的コントロールとシェアストレス負荷を行った内皮細胞の核抽出液を用いたゲルシフトアッセイおよびスーパーシフトアッセイを行った。その結果、TRE、CRE、kappaBに結合する蛋白がシェアストレス負荷後1時間で最高に増加し、その後漸減した。スーパーシフトアッセイの結果から、TREと相互作用する主な転写制御因子としては、c-Jun、JunDのホモダイマーまたはc-Jun、JunDとc-Fos、Fra1、Fra2のヘテロダイマーとなるいわゆるAP-1と考えられた。CREについてはCREB-1及びCREM-1のホモまたはヘテロダイマーと考えられた。kappaBについては、p50及びp52のホモまたはヘテロダイマーからなるNF-kappaBと考えられた。 4)シェアストレス情報伝達機構に関与する情報伝達因子を特定する為に、シェアストレスによるTRE、CRE、kappaB転写誘導について、それぞれ各種阻害剤を用いて検討した。その結果、TREについてはチロシンキナーゼ阻害剤・HerbimycinAが最も顕著な阻害効果を有し、次いでNO産生阻害剤・L-NMMAが有意に阻害効果を示した。MAPキナーゼ阻害剤・PD98059およびSB202190については後者が有意な阻害効果を示し、p38MAPkinaseの関与を示唆した。Srcファミリー特異的チロシンキナーゼ阻害剤・PP2は有意な阻害効果は無かった。SODは効果が無かったがcatalaseは部分的阻害効果を示し、ROSの関与を示唆した。一方、Ca2+キレート剤BAPTA-AM、G-protein Ras阻害剤・B581は効果が認められなかった。CREについてはHerbimycinAおよびBAPTA-AMが最も有意な阻害効果を示し、次いでPKA阻害剤・H-89が阻害効果を示した。kappaBについてはHerbimycinA、BAPTA-AM、L-NMMAの順に有意な阻害効果を示した。以上の結果から、三つのシスエレメントについて共通に阻害効果の認められたチロシンキナーゼが、シェアストレス情報伝達機構の上流側に存在すると考えられる。 5)チロシンキナーゼがシェアストレス情報伝達機構の上流に位置する可能性が示されたことから、シェアストレスによる細胞内チロシンキナーゼ活性の時間変化を検討した。シェアストレス15dyne/cm2の流れ負荷を0、1、3、5、10、15、30、60、120分間、BFAEに負荷し、それぞれ細胞溶解液を作成した。これを抗リン酸化チロシン抗体で免疫沈降し、得られた免疫沈降物でSDS-PAGEを行い、抗リン酸化チロシン抗体でウェスタンブロットを行った。その結果、30KD、45-65KD、120KD付近にシェアストレス依存的にチロシンリン酸化を受けるタンパク質を認めた。チロシンリン酸化は、シェアストレス負荷後1分以内に上昇し始め、10分後に極大に達した。15分後には減少に転ずるが、その後もbasal levelより若干高い状態を維持した。 6)接着斑(focal contact)は力学的刺激受容装置の一つと目されている。これに基づき、接着班に局在することが知られるFAKのシェアストレスによる活性化を検討した。その結果、FAKはシェアストレス負荷後10分でチロシンリン酸化が極大となることを見出した。これは、FAKがシェアストレス依存的に活性化されておりシェアストレス情報伝達に関与している可能性が示唆された。 7)FAKのシェアストレス情報伝達機構への関与を検討する為、TREルシフェラーゼリポーターと、FAK、FRNK、ミュータントFAK発現ベクター又はコントロール用発現ベクターをコトランスフェクションして、シェアストレス依存的転写誘導能を検討した。その結果、コントロールでは約15倍の転写誘導を認めたのに対し、FAK強制発現系では約17倍の転写誘導を示し、阻害効果はなかった。ミュータントFAK・K454R(キナーゼ活性欠損)、Y397F(自己リン酸化部位変異)、Y925F(Srcリン酸化部位変異)強制発現系においても阻害効果は認められなかった。一方、FRNK強制発現系では約4倍の転写誘導に止まり、約70%の阻害効果を認めた。上記の結果から、FAKが正常な機能を発揮する際に立体構造が正常に保たれていることが重要であることが示唆される。以上の結果は、シェアストレス情報伝達機構にFAKが関与することを強く示唆するものである。 以上本論文は、流れ負荷によるシェアストレス応答性転写因子群(AP-1、CREB-1及びCREM-1、NF-kappaB)を介した遺伝子発現の情報伝達にはチロシンキナーゼが共通して関与しており様々な経路が活性化されるシェアストレス情報伝達経路においてチロシンキナーゼが上流側に存在することを示し、さらに、力学的刺激感知部位の一つと考えられている接着斑に存在するFAKがシェアストレス情報伝達に関与するチロシンキナーゼであることを初めて明らかにしたものであり、学位の授与に値するものと考えられる。 | 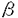 等の増殖因子の産生亢進、ICAM-1やVCAM-1等の接着分子の発現亢進あるいは減少などの反応を起こす。さらにシェアストレスで内皮機能が変化する際、その機能に関連した遺伝子の発現も変化することが多い。これまで、シェアストレスによる遺伝子発現調節に転写を介する機序とmRNAの安定化を介する機序があること、また、前者に於いてはシェアストレスにより活性化を受ける転写因子とそれが結合する遺伝子プロモータ上のシス・エレメントすなわちシェアストレス応答配列の幾つかが明らかされている。しかし、現在のところ、シェアストレスによる遺伝子転写調節に関わる情報伝達因子については検索がまだ十分進んでいない。こうした問題を解明することは力学的刺激受容体としての血管内皮細胞の性質を明らかにするだけでなく、血流依存性に生じる現象とされている血管新生、血管のリモデリング、ヒトの粥状動脈硬化症などの発生機序の理解に寄与するところが極めて大きいと思われる。そこで本研究では、血管内皮細胞におけるシェアストレス応答性転写因子群(AP-1、CREB-1及びCREM-1、NF-kappaB)を介した遺伝子発現誘導に関与する細胞内情報伝達機構を解析した。その結果、流れ負荷による遺伝子発現誘導がシェアストレス依存性であること、チロシンキナーゼがシェアストレス応答性転写因子群を介した転写誘導に共通する情報伝達因子であること、さらにFAK(Focal Adhesion Kinase)がシェアストレス依存的遺伝子発現誘導に関与することを示す結果を得たのでここに報告する。
等の増殖因子の産生亢進、ICAM-1やVCAM-1等の接着分子の発現亢進あるいは減少などの反応を起こす。さらにシェアストレスで内皮機能が変化する際、その機能に関連した遺伝子の発現も変化することが多い。これまで、シェアストレスによる遺伝子発現調節に転写を介する機序とmRNAの安定化を介する機序があること、また、前者に於いてはシェアストレスにより活性化を受ける転写因子とそれが結合する遺伝子プロモータ上のシス・エレメントすなわちシェアストレス応答配列の幾つかが明らかされている。しかし、現在のところ、シェアストレスによる遺伝子転写調節に関わる情報伝達因子については検索がまだ十分進んでいない。こうした問題を解明することは力学的刺激受容体としての血管内皮細胞の性質を明らかにするだけでなく、血流依存性に生じる現象とされている血管新生、血管のリモデリング、ヒトの粥状動脈硬化症などの発生機序の理解に寄与するところが極めて大きいと思われる。そこで本研究では、血管内皮細胞におけるシェアストレス応答性転写因子群(AP-1、CREB-1及びCREM-1、NF-kappaB)を介した遺伝子発現誘導に関与する細胞内情報伝達機構を解析した。その結果、流れ負荷による遺伝子発現誘導がシェアストレス依存性であること、チロシンキナーゼがシェアストレス応答性転写因子群を介した転写誘導に共通する情報伝達因子であること、さらにFAK(Focal Adhesion Kinase)がシェアストレス依存的遺伝子発現誘導に関与することを示す結果を得たのでここに報告する。