【序】カルシウムは、生体内の様々な刺激応答反応において、最も重要かつ普遍的なシグナル物質として働いている。これらCa2+の作用は、Ca2+結合タンパク質を介して発揮される。Ca2+結合タンパク質は非常に多岐に渡るが、中でもEF-handタンパク質は、Ca2+シグナルのトランスデューサーとして、主要な役割を担うものが多い。 40kDa Calcium-binding protein(CBP40)は真性粘菌の変形体特異的に発現しているEF-handタンパク質である。細胞内の主要なタンパク質の一つであるが、細胞外にも存在する。変形体細胞の損傷に応答して、膜の修復に機能すると考えられている。アミノ酸355残基から成り、C末端領域に4つのEF-hand配列を含む。変形体細胞においては、CBP40の一部はプロテアーゼによって分解され、N末端32残基を欠失した N32CBP40となる。CBP40はCa2+を結合すると凝集して沈殿する性質があるが、N32CBP40はCa2+を結合しても会合しない。 N32CBP40となる。CBP40はCa2+を結合すると凝集して沈殿する性質があるが、N32CBP40はCa2+を結合しても会合しない。 現在までに多くのEF-handタンパク質の立体構造が報告されている。しかしながら、それらの大部分がEF-handドメイン単独の構造であり、マルチドメインのEF-handタンパク質において、EF-hand構造が他のドメインとどのように相互作用し、機能に関わっているかについての情報はほとんど無い。本研究では、マルチドメインのEF-handタンパク質N32CBP40のCa2+遊離型・結合型の立体構造をX線結晶解析によって決定し、EF-handと他のドメインの相互作用について、また、Ca2+による調節機構、Ca2+高親和性について考察した。 【実験】大腸菌の発現系より得られたN32CBP40をEDTA共存下で精製・結晶化し、Ca2+遊離型の結晶を得た。並行してCa2+共存条件でCa2+結合型の結晶化条件を探索したが、再現性良く結晶を得ることは困難であった。そこで、Ca2+遊離型の結晶を塩化カルシウム溶液中にソーキングすることにより、Ca2+結合型の結晶を得た。Ca2+遊離型・結合型結晶は共に分解能3.0Åで、溶媒含有率は63%と非常に高かった。分解能の向上を目指して、精製度の向上・界面活性剤や添加剤などの検討・低温測定・架橋2量体や変異体の結晶化などを試みたが、効果は得られなかった。 水銀・サマリウムの重原子誘導体を作成し、多重重原子同型置換法によりCa2+遊離型の構造を決定した。得られたCa2+遊離型の構造をプローブとし、分子置換法によりCa2+結合型の構造を決定した。 Ca2+結合型構造では、EF1-4上にCa2+の電子密度が明瞭に認められ、Ca2+が4個結合していることが確認された。一方Ca2+遊離型では、Ca2+高親和性のため、残存Ca2+の電子密度が、弱いながらも認められた。残存Ca2+の電子密度はEF2,EF3において特に弱かったことから、EF2,EF3はCa2+低親和性サイト、EF1,EF4はCa2+高親和性サイトと考えられた。誘導プラズマ原子発光分析を行い、残存Ca2+の占有率を定量したところ、各Ca2+結合部位につき2-5%にすぎないことが明らかになった。以上より、得られた構造が、それぞれCa2+結合型・遊離型であることが確認された。 【結果と考察】全体構造は、Ca2+遊離型・結合型共に、coiled-coil、ヘリックスリッチ、EF-handの3つのドメインから構成されており、ドメイン間の相互作用面積は非常に広かった(図1)。このようにEF-handドメインが他のドメインと一体となった構造は、N32CBP40が初めての例である。 N32CBP40はカルモジュリンの10倍以上高いCa2+親和性を示すが、coiled-coil・ヘリックスリッチドメインを欠失させると、Ca2+親和性が低下する。coiled coil・ヘリックスリッチドメインは、EF-handドメインと広い面積に渡って相互作用することにより、EF-handドメインがCa2+を強く結合するのに適した構造を安定に保持する役割を担っているのであろう。 coiled-coilドメインは2本の長い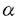 -ヘリックスから成り、ヘリックスリッチ・ EF-handドメインの周囲に沿って走っていた。このような球状構造の一部を構成するcoiled-coil構造は、既知のcoiled-coil構造のいずれにも該当せず、N32CBP40独自のユニークな構造として注目される。 -ヘリックスから成り、ヘリックスリッチ・ EF-handドメインの周囲に沿って走っていた。このような球状構造の一部を構成するcoiled-coil構造は、既知のcoiled-coil構造のいずれにも該当せず、N32CBP40独自のユニークな構造として注目される。 ヘリックスリッチドメインは、6本のヘリックスから構成される。溶媒中に露出されたループ上に、細胞接着活性を持つコンセンサス配列として知られるRGD配列が存在する(図1)。このループ構造は、細胞外接着タンパク質におけるRGD配列の構造と共通しているが、実際に機能を持つかどうかは、現在のところ不明である。  図1 Ca2結合型の全体構造のステレオ図。Ca2+を球で示す。 図1 Ca2結合型の全体構造のステレオ図。Ca2+を球で示す。 EF-handドメインのトポロジーは、カルパインのドメインIV’に良く似ていた。両者とも、カルモジュリンとは異なってEF1/2とEF3/4の間のループがないため、EF1/2とEF3/4が独立に運動できなくなっているのが特徴である。従来、CBP40とカルパインは、早くから分岐した別の進化経路を経てきたと考えられていたが、トポロジーが類似していたことから、遺伝子重複や融合の過程は似ていたものと思われる。 Ca2+遊離型・結合型構造を比較したところ、EF-handループ側鎖のコンフォメーション以外は、相違はほとんど見られなかった。Ca2+遊離状態においても、既にCa2+結合状態の構造がほとんど完成されていることに注目される。Ca2+結合に伴う構造変化が小さい理由として、CBP40はEF1/2とEF3/4の間のループが無く、両者が相互作用して安定な球状構造を作っているため、大きな構造変化が起こりづらいという点が挙げられる。Ca2+結合型構造は遊離型とほとんど同じであったにも関わらず、Ca2+との共結晶化が困難であったことから、Ca2+結合型は複数のコンフォメーションを取り、今回得られた構造はそのうちの一つである可能性も考えられる。 構造変化はほとんどなかったにも関わらず、Ca2+の結合に伴って、EF-handループを中心に温度因子の大幅な低下が観察された。これは、Ca2+を結合するとEF-handループの運動性が低下し、構造が堅固になることを示唆する。Ca2+結合に伴う温度因子の低下は,EF-handのみならず、coiled-coil・ヘリックスリッチドメイン中の残基でも認められた。これらの残基は、EF-handドメインとの相互作用面を中心として、その周囲に広がって分布していた(図2)。この分布から,Ca2+結合によって引き起こされたEF-handループの運動性の低下が、ドメイン間接触面を経由して、coiled coil・ヘリックスリッチドメインにまで広範囲に渡って伝達されると考えられる。この変化は裏側の分子表面残基にまで伝わっており、自己会合や標的との相互作用に、影響を与える可能性も考えられる。  図2 (左)ドメイン間相互作用面。coiled-coil・ヘリックスリッチドメイン上の残基で、EF-handドメインと相互作用しているものを淡灰色で示す。 (右)左図と同じ方向から見た図。Ca2+結合に伴って,温度因子が低下した残基を濃灰色で示す。 図2 (左)ドメイン間相互作用面。coiled-coil・ヘリックスリッチドメイン上の残基で、EF-handドメインと相互作用しているものを淡灰色で示す。 (右)左図と同じ方向から見た図。Ca2+結合に伴って,温度因子が低下した残基を濃灰色で示す。 EF-handドメインは、coiled-coil・ヘリックスリッチドメインに対し、van der Waals結合と疎水性相互作用によって相互作用していた。van der Waals結合は非共有結合の中で最も弱い結合であり、また、疎水性相互作用は結合原子間の方向性に制限がないという特徴を持つ。これらの性質は、運動性の低下というシグナルを、広い範囲に渡って中断することなく伝達するのに適しているのかもしれない。 真核生物におけるCa2+の濃度分布は、細胞内で10-7〜10-8M、外的刺激を受けると10-5M程度まで上昇する。一方、細胞外では10-3M程度である。N32CBP40は、Kdが M以下のCa2+高親和性サイトと、M以上の低親和性サイトを2つずつ持つ。前述した電子密度の比較より、EF1,4がCa2+高親和性サイト、EF2,3が低親和性サイトに相当することが示唆された。これらのことから、静状態の細胞内においては、CBP40はEF1,4にCa2+を結合した状態であるが、刺激を受けると(あるいは細胞外に分泌されると)EF2,3にCa2+が結合し、活性状態になると考えられる。なお、CBP40のCa2+依存的な自己会合は、高親和性サイトにCa2+が結合しただけでは起こらず、低親和性サイトにCa2+が結合したときに初めて起こることが知られている。 M以下のCa2+高親和性サイトと、M以上の低親和性サイトを2つずつ持つ。前述した電子密度の比較より、EF1,4がCa2+高親和性サイト、EF2,3が低親和性サイトに相当することが示唆された。これらのことから、静状態の細胞内においては、CBP40はEF1,4にCa2+を結合した状態であるが、刺激を受けると(あるいは細胞外に分泌されると)EF2,3にCa2+が結合し、活性状態になると考えられる。なお、CBP40のCa2+依存的な自己会合は、高親和性サイトにCa2+が結合しただけでは起こらず、低親和性サイトにCa2+が結合したときに初めて起こることが知られている。 4つのEF-handの中でもEF2,3は、coiled-coil・ヘリックスリッチドメインとの非共有結合の数が多かった。これは、EF1,4が閉構造をとって疎水性残基をEF-hand内部に収めているのに対し、EF2,3は構造が「開いて」おり、疎水性残基が露出しているからである。coiled-coil・ヘリックスリッチドメインにおいて温度因子が低下した残基は、EF2,3との相互作用部位付近に特に多く見られた。 以上より、Ca2+低親和性サイトであるEF2,3は、Ca2+が結合したという情報をcoiled-coil・ヘリックスリッチドメインに伝え、機能調節を行うのに最も重要な部位であると考えられる。一方、Ca2+高親和性サイトであるEF1,4は、EF2,3とそれぞれ対を形成することにより、立体構造の安定化の役割を担っていると考えられる。 |