学位論文要旨
| No | 115042 | |
| 著者(漢字) | 仲村,厚志 | |
| 著者(英字) | Nakamura,Atsushi | |
| 著者(カナ) | ナカムラ,アツシ | |
| 標題(和) | ニワトリ松果体光受容タンパク質ピノプシンの機能解析 | |
| 標題(洋) | Functional analysis of chicken pineal photoreceptive molecule pinopsin | |
| 報告番号 | 115042 | |
| 報告番号 | 甲15042 | |
| 学位授与日 | 2000.03.29 | |
| 学位種別 | 課程博士 | |
| 学位種類 | 博士(理学) | |
| 学位記番号 | 博理第3806号 | |
| 研究科 | 理学系研究科 | |
| 専攻 | 生物化学専攻 | |
| 論文審査委員 | ||
| 内容要旨 | ニワトリ松果体は光受容能を持ち、受容された光シグナルは松果体細胞に内在する概日時計の位相を調節する。ニワトリ松果体の光受容タンパク質ピノプシンは、網膜以外の組織から初めてクローニングされたオプシンであり、アミノ酸配列を基に作成した分子系統樹において、網膜の光受容タンパク質(視物質)とは異なる新しいグループを形成する。ピノプシンのクローニング以降、さらにいくつかの光受容タンパク質が網膜外組織からクローニングされ、概日時計の光位相同調や日長識別など多様な光生理現象に関与すると想定されてきた。しかしながら、視書質の性状解析に関しての膨大な研究がある一方で、網膜外の光受容タンパク質の性質については、試料の調製が困難なことから、そのほとんどが謎に包まれている。そこで本研究では、まず培養細胞でのピノプシンの大量発現系および精製方法を確立した。次に、精製したピノプシンを用いて、光吸収後の構造変化過程とGタンパク質との共役過程を、分光学的・生化学的手法を駆使して詳細に検討した。 ピノプシンは、松果体に極く微量しか存在せず(2ng/松果体)、機能解析を行うために必要な量のピノプシンを精製することは極めて困難である。そこで、アフィニティー精製のためのタグ(6残基のヒスチジン)をN末端に導入したピノプシンを設計し、培養細胞293Sを用いて強制発現させた。発現したピノプシン(アポ蛋白質)に11シス型レチナールを加えて光受容タンパク質を再構成したのち、界面活性剤Dodecyl maltosideを用いて可溶化した。この細胞抽出液を出発材料として、3段階のカラムクロマトグラフィー(DEAE-Sepharoseカラム、ニケッルキレートカラムおよびSP-Sepharoseカラム)を行うことにより、ピノプシンを高純度に精製することに成功した。また、ヒスチジンタグを付加したニワトリロドプシン(桿体視物質)とニワトリGreen(緑色感受性錐体視物質)を同様に発現させて精製し、ピノプシンと共に以下の測定に用いた。 まず、低温吸収スペクトル法によりピノプシンの光退色過程を調べた。その結果、ピノプシン(吸収極大波長:468nm)は視物質と同様、光を吸収した後、バソ(527nm)、ルミ(461nm)、メタI(460nm)、メタII(385nm)、そしてメタIII(460nm)という一連の中間体を経て退色した。この退色中間体系列とその分光学的性質は視物質のそれらと良く一致することから、ピノプシンの光受容後の構造変化は視物質と類似していることが示唆された。 次に、ウシ網膜より精製したGタンパク質トランスデューシン(Gt)を用いてGTP 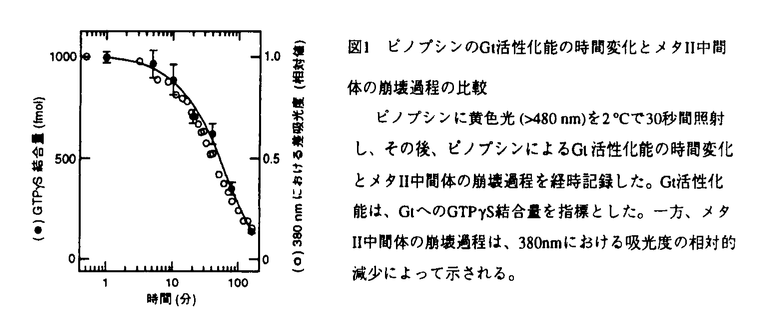  S結合量を指標とした。一方、メタII中間体の崩壊過程は、38nmにおける吸光度の相対的減少によって示される。 S結合量を指標とした。一方、メタII中間体の崩壊過程は、38nmにおける吸光度の相対的減少によって示される。ロドプシンと錐体視物質のメタII中間体の性質は互いに大きく異なり、この違いが光受容細胞(桿体と錐体)の光応答特性の特徴を生み出す一因と考えられている。また、アミノ酸配列の比較から、ピノプシンはロドプシンよりも錐体視物質に類似した性質を持つと推定される。そこで、ピノプシンのメタII中間体の性質(生成および崩壊の速度)をロドプシンおよびGreenと比較した(図2、上段)。その結果、ピノプシンのメタII中間体の生成はロドプシンよりも20倍以上速く(-25℃における時定数6.2分)、Greenとほぼ同じ速度で生成した(つまり錐体タイプ)。一方、メタII中間体の崩壊過程は(図2、下段)、予想に反してピノプシンはGreenより80倍以上遅く、すなわちロドプシンとほぼ同じ速度(2℃における時定数44分)でゆっくりと崩壊した(つまり桿体タイプ)。これらの結果より、ピノプシンのメタII中間体は桿体と錐体の両方の視物質の性質を兼ね備えていることが判明し、ピノプシンは新しいタイプの光受容タンパク質であることが明らかになった。ピノプシンのメタII中間体が予想以上に安定であることから、松果体における光情報伝達の過程では、網膜よりも時間分解能は低いものの光情報をより高度に増幅できる可能性が考えられる。このことは、脳における概日時計の光同調というピノプシンの生理機能に適した特性といえるかもしれない。また、ピノプシンとロドプシン・錐体視物質の系統関係を考え併せると、ピノプシンはメタII中間体の寿命を延ばす機能を、ロドプシンとは独立に獲得したことが示唆される。  そこで次に、ピノプシンのメタII中間体が錐体視物質よりも強く安定化される機構をアミノ酸レベルで明らかにするために、部位特異的な変異を導入した変異ピノプシンを用いた解析を行った。まず、候補となるアミノ酸残基を絞り込むために、既知の錐体視物質において保存されており、かつピノプシンにおいては異なるアミノ酸残基を検索した。この基準を満たす部位は、(i)ピノプシンの171番目のSer、(ii)184番目のAsn、(iii)2残基の欠失(190番目と191番目の間)、の3カ所であった。ピノプシンのこれらの部位を錐体視物質型のアミノ酸残基に置換した変異体を作成し、メタII中間体の崩壊過程を測定した(図3)。その結果、171番目および184番目それぞれの残基を置換した変異体P-S171RおよびP-N184Dでは、メタII中間体の寿命はそれぞれピノプシンの約70%および約80%となったが、両方の残基の2重変異体P-S171R/N184Dではピノプシンとほぼ同じ寿命を示した(図3、右図)。このことから、これら2残基(の組み合わせ)はピノプシンのメタII中間体の安定化にほとんど寄与していないと考えられる。一方、2残基を挿入した変異体P-188Gr193は11シス型レチナールと結合しなかったが、3つの部位を全て置換した変異体P-S171R/N184D/188Gr193では、メタII中間体の寿命がピノプシンの約30%まで減少した(図3、右図)。以上の結果より、190番目と191番目の間の2残基の欠失が、ピノプシンのメタII中間体の安定性に重要な役割をはたしていることが明らかになった。この2残基の欠失はピノプシンの細胞外側の第2ループにみられ、桿体視物質のロドプシンでは、この領域がタンパク賀全体の正常なフォールディングに大きな影響を与えることが知られている。私の得た結果は、ピノプシンにおいてもこの領域がタンパク質の高次構造の形成、ならびにメタII中間体の安定性の制御に関わっていることを示している。 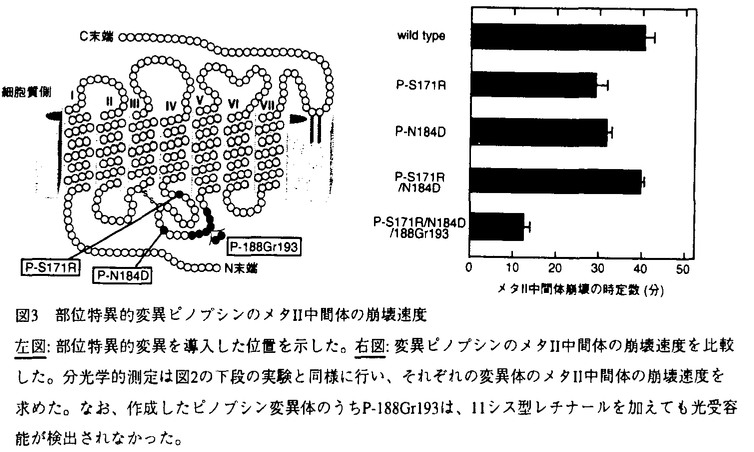 | |
| 審査要旨 | 本論文で扱われたニワトリ松果体の光受容タンパク質ピノプシンは、網膜以外の組織から初めてクローニングされたオプシンであり、概日時計の光位相同調に関わると考えられる。ピノプシンのクローニング以降、さらにいくつかの光受容タンパク質が網膜外組織からクローニングされ、概日時計の光位相同調や日長識別など多様な光生理現象に関与すると想定されてきた。しかしながら、網膜の光受容タンパク質(視物質)の性状解析に関しての膨大な研究がある一方で、網膜外の光受容タンパク質の性質については、試料の調製が困難なことから、そのほとんどが謎に包まれている。そこで本研究では、ピノプシシの機能解析を行うために、まず培養細胞でのピノプシンの大量発現系および精製方法を確立した。 ピノプシシは、松果体に極く微量しか存在せず(2ng/松果体)、機能解析を行うために必要な量のピノプシンを精製することは極めて困難である。そこで、アフィニティー精製のためのヒスチジンタグを導入できるようにピノプシンcDNAを改変し、培養細胞293Sを用いて強制発現させた。発現したピノプシンに11シス型レチナールを加えて光受容タンパク質を再構成したのち、界面活性剤Dodecyl maltosideを用いて可溶化した。この細胞抽出液を出発材料として、3段階のカラムクロマトグラフィーを行うことにより、ピノプシンを高純度に精製することに成功した。精製したピノプシン試料を用いて以下の解析を行った。 まず、低温吸収スペクトル法によりピノプシンの光退色過程を調べた。その結果、ピノプシンは視物質と同様、光を吸収した後、バソ、ルミ、メタI、メタII、そしてメタIIIという一連の中間体を経て退色した。この退色中間体系列とその分光学的性質は視物質のそれらと良く一致することから、ピノプシンの光受容後の構造変化は視物質と類似していることが示唆された。 ロドプシンと錐体視物質のメタII中間体の性質は互いに大きく異なり、この違いが光受容細胞(桿体と錐体)の光応答特性の特徴を生み出す一因と考えられている。また、アミノ酸配列の比較から、ピノプシンはロドプシンよりも錐体視物質に類似した性質を持つと推定される。そこで、ピノプシンのメタII中間体の性質をロドプシンおよび緑色感受性の錐体視物質Greenと比較した。その結果、ピノプシンのメタII中間体の生成はロドプシンよりも20倍以上速く、Greenとほぼ同じ速度で生成した(錐体タイプ)。一方、メタII中間体の崩壊過程は、予想に反してピノプシンはCreenより80倍以上遅く、ロドプシンとほぼ同じ速度でゆっくりと崩壊した(桿体タイプ)。これらの結果より、ピノプシンのメタII中間体は桿体と錐体の両方の視物質の性質を兼ね備えていることが判明し、ピノプシンは新しいタイプの光受容タンパク質てあることが明らかになった。 ピノプシンのメタII中間体が錐体視物質よりも強く安定化される機構をアミノ酸レベルで明らかにするために、部位特異的な変異を導入した変異ピノプシンを用いた解析を行った。まず、候補となるアミノ酸残基を絞り込むために、既知の錐体視物質において保存されており、かつピノプシンにおいては異なるアミノ酸残基を検索した。この基準を満たす部位は、(i)ピノプシシの171番目のSer、(ii)184番目のAsn、(iii)2残基の欠失(190番目と191番目の間)、の3カ所であった。ピノプシンのこれらの部位を錐体視物質型のアミノ酸残基に置換した変異体を作成し、メタII中間体の崩壊過程を測定した。その結果、171番目と184番目の両方の残基を置換した変異体では、メタII中間体の寿命は野生型とほぼ同じ値を示した。2残基を挿入した変異体は11シス型レチナールと結合しなかったが、3つの部位を全て置換した変異体では、メタII中間体の寿命が野生型の約30%まで減少した。以上の結果より、190番目と191番目の間の2残基の欠失が、ピノプシンのメタII中間体の安定性に重要な役割をはたしていることが明らかになった。この欠失は他の全ての網膜外光受容タンパク質にも見られることから、これらのタンパク質のメタII中間体も安定化されていることが予想される。このことから、網膜外組織における多様な光生理現象には、光情報の増幅が必要であることが推測される。 なお、本論文は、小島大輔、岡野俊行、今井啓雑、寺北明久、七田芳則、深田吉孝氏との共同研究であるが、論文提出者が主体となって研究を行ったものであり、論文提出者の寄与が充分であると判断できる。 したがって、博士(理学)の学位が授与できると認める。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/54778 |