本論文は2章からなり、第1章は、"Identification and characterization of short inflorescence,early senescence mutant,fireworks(fiw)"(訳:花茎の短い突然変異体fireworksの単離・同定とその特徴)について、第2章は、"Identification and characterization of brassinosteroid-related dwarf mutants,chibi(chi)"(訳:ブラシノステロイド関連矮性突然変異株chibiの単離・同定とその特徴)について、述べられている。 本研究では、モデル植物シロイヌナズナを用いて、植物の伸長成長に関する新奇の突然変異体を単離し、その解析により、植物の伸長成長に関する遺伝的プログラムのより深い理解を行なうべく研究が行われた。まず、形質転換シロイヌナズナの作出にあたり、減圧浸潤法の改良により形質転換効率の向上を図り、T-DNA挿入形質転換体約2500ヶを選抜した。得られた形質転換体は、カリフラワーモザイクウイルス35Sプロモーターの転写エンハンサーを4コピー含んでいるので、機能欠失型・獲得型の双方の変異体の出現が期待された。 第一の突然変異体は、花茎特異的な伸長抑制を示す変異体を同定したが、その形態が花火に似ていることからfireworks(fiw)と名付けたが、この変異体は栄養成長期には特別な変異は見られず、特定の生育段階でのみ矮性を示した。遺伝学的解析から、fiwは挿入T-DNAと変異は連鎖しない劣性変異であり、その染色体上での変異の場所は、第44番染色体の下方の約448kbpの領域内であると同定した。続いて、fiwで見られた花茎の伸長抑制のメカニズムを探究したが、表皮細胞ではfiw個体の矮性の度合いとほぼ相当する細胞の伸長抑制が認められたが、ジベレリン投与によってはfiwの表現型は回復しなかった。従って、fiwの花茎が示す矮性は細胞伸長の欠損に由来するが、ジベレリン合成欠損がその原因ではないと判断した。 第二の突然変異体は、胚軸の光シグナルの応答に関するもので、結果的にブラシノステロイド関連の変異体であった。作出されたタグラインから、弱い近赤外光下でより強い胚軸伸長抑制を示す三系統の変異体を得、これらにchibi 1、2、3と名付けた。これらは成熟個体でも強い矮性を示し、chi2は優性、それ以外は劣性の突然変異であった。優性変異は、過剰発現型ベクターによって本来期待され変異であるので、chi2変異に関して詳細に解析したところ、変異は第1番染色体上方にマップされ、chi2のT-DNA挿入位置の近傍に新奇のシトクロムP450遺伝子(CYP72C1)を見出された。この遺伝子がchi2変異体では過剰発現されていることを確認するとともに、CYP72C1を過剰発現する形質転換植物を作製することによっても、chi2変異体と同様の表現型を観察した。これらにより、CYP72C1がchi2変異の原因遺伝子と結論した。なお、chi2変異体の形態は、既知のブラシノステロイド欠損突然変異体と極めて類似しているので、ブラシノステロイドの添加実験を行ったところ、chibi2変異体芽生えの伸長抑制は濃度依存的に回復した。また、ブラシノステロイド合成の中間代謝産物の内生含量は、6-deoxotyphasterol(typhasterol)とその下流でchi2において極端に減少していた。これらの結果は、CYP72C1遺伝子産物が本来の合成反応とは異なる反応を触媒した為と考えられる。 従って、本研究の二種類の突然変異体は新奇のものであり、いずれも植物の伸長成長の遺伝プログラムに関した新知見をもたらすものであると判断される。 なお、本論文の第1章は、長谷あきら氏、望月伸悦氏との共同研究であるが、論文提出者が主体となって研究のデザインを計画し、遂行したので、論文提出者の寄与が十分であると評価する。 よって、博士(理学)の学位を授与できると認める。 | 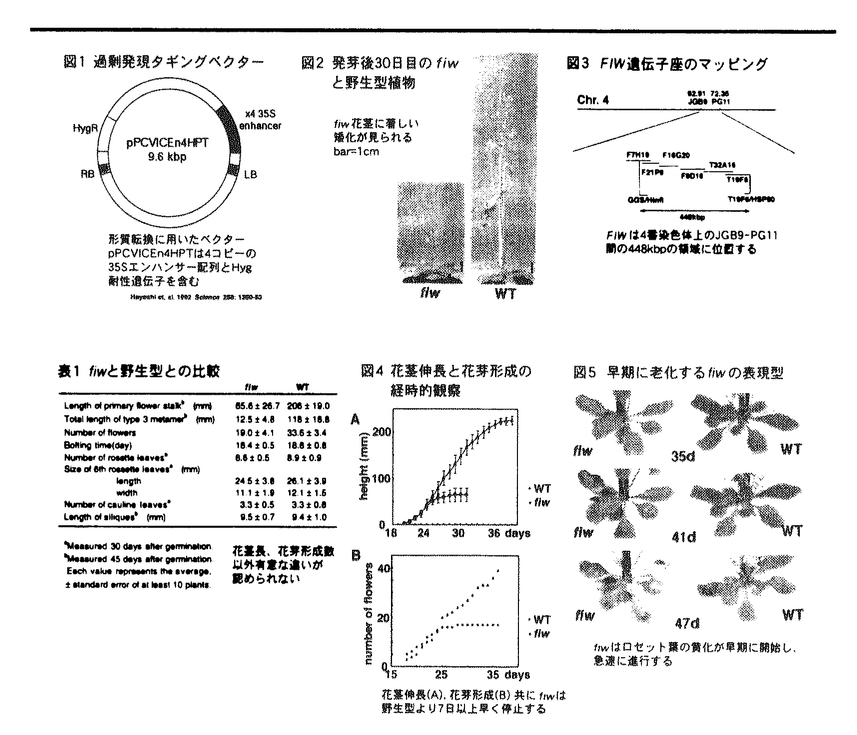
 Mのブラシノステロイド存在下で完全に回復した(図11)。一方同様の方法でジベレリンを添加したところ、ジベレリンは胚軸伸長に影響を与えなかった。続いてブラシノステロイド合成の中間代謝産物の含量を測定したところ、6-deoxotyphasterol(typhasterol)とその下流の代謝産物がchi2において極端に減少していた。
Mのブラシノステロイド存在下で完全に回復した(図11)。一方同様の方法でジベレリンを添加したところ、ジベレリンは胚軸伸長に影響を与えなかった。続いてブラシノステロイド合成の中間代謝産物の含量を測定したところ、6-deoxotyphasterol(typhasterol)とその下流の代謝産物がchi2において極端に減少していた。