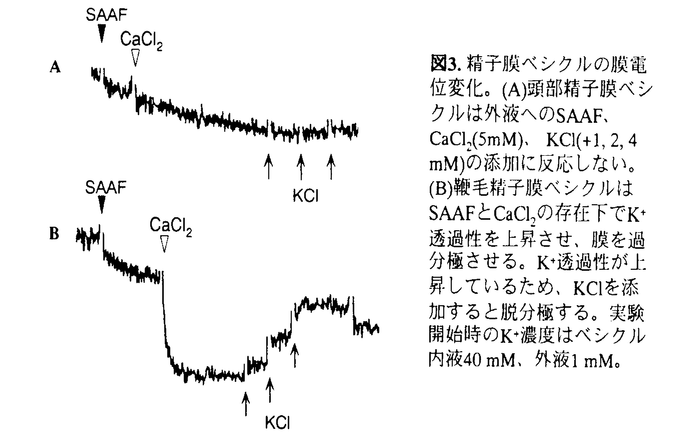
本論文は3章からなり、第1章は、原索動物ホヤの精子運動活性化にK+チャネルの活性化及び細胞膜の過分極が必須であること、第2章は、ホヤ精子において細胞膜の過分極がadenylyl cyclaseを活性化し、その結果合成されるcAMPが精子運動活性化の引金を引くこと、第3章は、ホヤ精子細胞膜のion channelsの解析について述べられている。 カタユウレイボヤCiona intestinalisとユウレイボヤC.savignyiの精子は海水中で運動を停止しており、卵由来精子活性化誘引物質(sperm-activation and-attracting factor:SAAF)の作用により精子内のcAMPを合成し、それが精子の運動を活性化することが明らかにされていた。しかし、SAAFの細胞膜における細胞情報伝達の機構については不明であった。第1章ではK+ionophore、valinomycin(Val)がSAAFと同様精子運動を活性化すること、細胞外K+濃度の上昇が精子運動活性化を抑制することを示し、細胞膜のK+透過性が精子運動活性化に重要である事を示した。更に、K+透過性を強く反映する膜電位変化を蛍光指示薬DisC3(5)を用いて測定し、運動停止精子ではK+透過性が低く静止膜電位は-50mVであり、SAAFの添加によりK+透過性が急激に上昇し、細胞内K+濃度が減少すると共に細胞膜は大きく過分極して-100mVとなることが明らかになった。更に膜の過分極がK+チャネル阻害剤MCD-peptideで抑制されること、イオン選択性はK+≧Rb+≫Cs+>Li+≧Na+であることから、SAAFはK+チャネルを活性化し、K+流出による膜の過分極を引き起こし精子運動を活性化することが明らかにされた。 第2章では、第1章の知見に基づき、膜の過分極とcAMP合成について調べた。その結果、Valは単独でSAAFと同様精子のcAMPを上昇させた。この反応は細胞外Ca2+に依存しない。次に2mM IBMX処理で人為的に細胞内cAMPを増加させた精子にSAAF又はValを加えたところ、更にcAMP濃度の上昇が見られたことから、運動を停止している精子ではphosphodiesterase活性が高いためcAMP濃度が低いこと、SAAFは膜の過分極を引き起こしadenylyl cyclase活性を上昇させcAMP濃度を上昇させると考えられる。また、K+チャネル阻害剤MCD-peptideはSAAFによるcAMP上昇を抑えるが、ValによるcAMP上昇は阻害しないことから、SAAFによるcAMP合成はK+チャネルの活性化に依存していることも示された。精子のadenylyl cyclase活性にCa2+依存性が見られない。したがって、ホヤ精子ではSAAFの作用で細胞膜が過分極し、それが直接adenylyl cyclaseを直接活性化しcAMP濃度を上昇させ、精子を活性化すること、Ca2+はこの酵素の活性化に関与しないことが明らかになった。 第3章では、精子細胞膜を窒素キャビテーション法で単離し精子膜ベシクルを作成し、細胞膜以外の細胞構成成分が関与しない系で精子運動活性化に関与するion channelの解析を行った。まず精子膜ベシクルを用いてその膜電位変化を蛍光指示薬DisC3(5)で測定したところ、ベシクル内液のK+濃度を外液の40倍高くし、SAAFを作用させるとベシクル膜のK+透過性が上昇し膜を過分極すること、鞭毛より単離した膜ベシクルはSAAFに反応するが頭部由来の膜ベシクルは反応しないことが明らかとなった。また外液のCa2+はSAAFによるK+透過性上昇を増大させた。以上の結果よりSAAF依存性のK+チャネルは精子鞭毛に局在すること、細胞外Ca2+はSAAFによるK+チャネルの活性化を増大させることが示唆された。次に膜ベシクルを人工脂質二重膜に融合させ、個々のion channel活動を解析した。その結果SAAF非存在下で活動し、K+を透過する陽イオンチャネルが観察された。また人工脂質二重膜に融合した精子ion channelの幾つかが、SAAFにより活性化されることも明らかとなった。 以上、第1章、第2章、第3章より、ホヤの精子細胞膜において、卵由来精子活性化誘引物質SAAFは鞭毛に局在するK+チャネルを活性化し、K+流出による膜の過分極を引き起こす。この膜電位変化が直接adenylyl cyclaseを活性化し、その結果合成されたcAMPが鞭毛運動を活性化することが明らかになった。また細胞外Ca2+がSAAFによるK+チャネルの活性化を増大させている可能性、細胞内Ca2+はadenylyl cyclaseの活性化に重要な役割を果たしていない可能性が明らかとなった。 なお、本論文の1章と第2章はMarian、稲葉、岡、森沢、第3章は岡、森沢との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断する。 したがって、博士(理学)の学位を授与できると認める。 | 

 M IBMX(□)では上昇しないが2mM IBMX(■)により上昇する。2mM IBMX処理した精子にSAAF(●)またはvalinomycin(▲)を作用させると、更にcAMP濃度が上昇する。
M IBMX(□)では上昇しないが2mM IBMX(■)により上昇する。2mM IBMX処理した精子にSAAF(●)またはvalinomycin(▲)を作用させると、更にcAMP濃度が上昇する。