本論文は4章からなり、第1章は、イネの子葉鞘の中の緑化しない細胞に於ける老化進行の解析、第2章はイネの第二葉葉身に於ける老化進行の解析、第3章は子葉鞘の老化に関わるヌクレースの解析、及び第4章はカルシウム指示蛍光試薬を用いた、子葉鞘内の老化の伝播様式の解析について述べられている。 高等植物の葉の老化は、発生の最終段階として、予め細胞内にプログラムされているものと考えられている(Smart 1994)。しかし、その機構については未だ全く明らかになっていない。老化の機構を解明するためには、先ず、老化に際して起こる事柄を総合的且つ詳細に把握することか必要である。この考えに基づき、本論文ではイネの子葉鞘を主な材料として用い、その老化進行について形態学的な視点から解析を行った。材料として主に用いている子葉鞘は、生活環か短く、構造が単純である、などの本研究を行うのに適した特徴を持っている。論文提出者は、修士課程において、子葉鞘の緑化した細胞について組織・細胞レベルでの老化の進行を蛍光顕微鏡・電子顕微鏡を用いて解析し、各現象の起こる時期・速度は異なるものの、全ての緑化する葉肉細胞内で(1)葉緑体DNAの減少、(2)細胞核の凝縮、葉緑体内膜の分解、(3)葉緑体の分解、細胞核の分解、(4)細胞内構造の崩壊、細胞壁の変形、と言った一連の過程を経て、老化か進行することを明らかにした(Inada et al.1998a,b)。本論文では、修士論文の結果を発展させ、最初に上記の現象の一般牲についての検証を行い、次に老化の機構に関する解析を進めた。最終的には、子葉鞘の中の最初に老化する始源細胞を突き止め、子葉鞘内の組織レベル・細胞レベルでの老化の順番を明らかにした。 第1章は、子葉鞘の緑化しない葉肉細胞の老化進行について述べられている。子葉鞘の外表皮近くの2-3層の細胞では播種後3日目に葉緑体が発達し、緑化する。一方、内側の3-7層の葉肉細胞では2日目に多量のデンプンが蓄積するが、その後一日で完全に分解される。テクノビット7100切片を用いた蛍光顕微鏡観察の結果、これらの外層と内層の細胞分化は1日目から2日目にかけて急速に進行し、その後3日目から4日目の間に内表皮から3-4層目の細胞で急激な細胞死が進行することがわかった。この細胞死はその後両表皮に向かって進行し、6日目には内層は完全に枯死して細胞壁が激しく変形する。内層の中の老化の早い内表皮から3層目の細胞と、老化の遅れる内表皮から1層目の細胞のオルガネラの変化を観察した結果、全ての細胞で、老化のごく初期にプラスチドDNAの減少、ミトコンドリアDNAの減少が起こり、1層目の細胞では老化の後期に著しい細胞核の凝縮が見られた。一方、3層目の細胞に於いては、細胞核の凝縮が見られず、4日目に急激に全ての細胞内構造が崩壊することがわかった。この結果より、子葉鞘の内層の細胞では、2つの種類の老化があることが示唆された。 第2章では、更に普通葉である第二葉を用いて現象の一般性について解析を行っている。第二葉は播種後3日目以降急速に成長し、葉身は4日目にはほぼ完全に伸長を終えるが、その後葉鞘が伸長し、第二葉全体としては7日目まで伸長を続ける。葉身は、6日目に完全展開し、8日目には先端部から老化が始まる。老化は基部に向かって進行し、11日目には先端から1/3の部分が枯死する。この先端から基部への老化の勾配は、生化学的解析によっても示された。葉身の先端部・基部についての蛍光顕微鏡解析の結果、老化の初期に於ける葉緑体DNAの減少、それに続く細胞核の凝縮が見られ、子葉鞘で観察された老化の一形態が、第二葉に於いても保存されていることが明らかになった。老化に於ける各現象の起こる時期・速度は、子葉鞘と第二葉、第二葉の先端と基部で違いが見られ、この老化の進行が、細胞の位置する場所、環境に依存して変化することが強く示唆された。 第1章、第2章に述べたイネ葉の老化の進行には、オルガネラDNAの分解にそれぞれ特異的に働くヌクレースが、重要な役割を果たしていると考えられる。そこで第3章では、子葉鞘の老化に関わるヌクレースについて解析を行った。組織内の老化時期の違いを反映した解析を行う為、フィルム基質法(Daoust and Amano 1957)を、フィルムDNAの濃度・染色法を変えて高感度に改変し、子葉鞘切片内のヌクレース活性を検出することに成功した。解析の結果、細胞核が著しく凝縮し分解される時期の細胞で、特異的にCa2+/Mg2+要求性のヌクレース活性が上昇していることが明らかになった。更に生化学的手法によるこのヌクレース分子の同定を試み、幾つかのヌクレース分子を同定した。 第3章で述べたフィルム基質法による解析の結果から、Ca2+/Mg2+要求性のヌクレース上昇を示す細胞内で、これらの金属イオン濃度が上昇しているのではないかとの仮説を立てた。これを検証するため、カルシウム指示蛍光試薬を用いた解析を第4章で行った。蛍光試薬による反応は、4日目にヌクレース活性が見られた内層の部分で特異的に検出され、若い時期の子葉鞘及び老化した外層の細胞では検出されなかった。この指示薬による染色が認められた部分は、通気組織が形成される場所と一致することから、通気組織形成に至るまでの過程を更に詳細に観察し、その結果、最初にCa2+の染色を示す細胞を維管束の近くに同定した。このCa2+反応はその後葉の中央部へ向かって一細胞層で伝播してゆき、更に両側表皮に向かって伝播する。更にこの細胞の構造を電子顕微鏡により詳しく観察したところ、液胞膜の崩壊に伴い、細胞内構造が著しく膨潤し、細胞死に至っていることがわかった。また、同一個体の先端部と基部から調整した切片を、カルシウム指示蛍光試薬で染色し、これらの細胞死が先端部から基部へ向かって伝播してゆくことを明らかにした。 以上の結果から、子葉鞘の葉肉細胞の老化は、主に二つの形態を取ることが明らかになった。即ち、組織的には維管束に向かって進行してゆき、徐々に進行するオルガネラの分解を特徴とする自然老化と、維管束近辺の一細胞から始まって葉の中央部に向かって伝播してゆき、液胞の崩壊に伴う急激な細胞内構造の分解を特徴とする細胞死型の老化である。老化のごく初期に於けるプラスチドDNA・ミトコンドリアDNAの分解は、両方の老化で同様に見られることから、基本的な老化開始のプログラムは全細胞に共通に保存されており、老化後期の過程が細胞の位置する場所により異なっている、と考えられる。 以上のように、本論文は、イネの子葉鞘という材料を用いてその老化の開始と進行順序について詳細に解析し、今後老化の機構へと研究を進めてゆく上での重要な基礎を築いた。この研究を基に更に解析を進めることにより、老化の開始と進行の機構が明らかになってゆくことが期待される。 尚、本論文第2章は、酒井敦、黒岩晴子、黒岩常祥の共同研究であるが、論文提出者が主体となって解析を行ったもので、論文提出者の寄与が十分であると判断する。 従って、博士(理学)の学位を授与できると認める。 | 
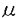 m / 図4 テクノビット7100切片によるオルガネラの経時的変化の観察 切片をDAPI、DiOC7で二重染色し、DNAと膜系の変化を調べた。内表皮から3層目の細胞(A)、1層目の細胞(B)の於ける細胞核(a)、プラスチドDNA(b)、ミトコンドリアDNA(b内部)、プラスチド膜(c)、ミトコンドリア膜(c内部)。写真左上の数字はそれぞれ播種後の日数を表す。バーは5
m / 図4 テクノビット7100切片によるオルガネラの経時的変化の観察 切片をDAPI、DiOC7で二重染色し、DNAと膜系の変化を調べた。内表皮から3層目の細胞(A)、1層目の細胞(B)の於ける細胞核(a)、プラスチドDNA(b)、ミトコンドリアDNA(b内部)、プラスチド膜(c)、ミトコンドリア膜(c内部)。写真左上の数字はそれぞれ播種後の日数を表す。バーは5