腸管は動物にとって栄養素吸収の最前線として重要な働きを担っている。栄養素吸収は腸管壁に一層に並んだ上皮細胞により行われるが、その吸収経路は大きく分けて4つに分けることが出来る。それは、1)グルコース、アミノ酸、ジペプチドなどがそれぞれに特異的な輸送担体によって輸送される経路、2)脂溶性ビタミン、脂肪酸などが特異的な結合タンパク質と結合し輸送される経路、3)タンパク質などの高分子がトランスサイトーシスにより輸送される経路、4)細胞同士の間隙を拡散により透過する経路である。このうち1)〜3)の経路については、関わる機能分子の特性や制御機構等が多くの研究者によって検討されてきた。一方、4)についてはその意義や調節の仕組みが不明であったため注目される事が少なかったが、近年、カルシウムなどの水溶性の低分子物質の透過経路としての重要性が認識されるようになってきた。細胞間には細胞同士を繋ぐ接着装置が備わっており、その中の1つであるタイト結合(TJ)が細胞間の物質透過性を制御するバリア機能を持つことも明らかになった。また、このTJはサイトカインや増殖因子などの内的因子、細菌毒素や食品成分などの外的因子により影響を受け、その結果、細胞間透過性を変化させる事が分かってきている。 一方、腸管には栄養素吸収のための上皮細胞のみならず、パイエル板、腸管膜リンパ節などの免疫担当組織が存在することも知られている。これらは腸管付属リンパ組織(GALT)と呼ばれ、外界からの異物に対する生体防御系として重要な役割を果たしていることから、近年、その機能は大きな注目を浴びるようになってきている。 このように腸管には栄養素吸収、生体防御と言う二つの大きな役割があるにも関わらず、この二つの関連についての情報は全く乏しいのが現状である。 以上の背景から、本論文では腸管上皮における物質吸収・透過性とサイトカインの関わりに着目し、1)機能性トリペプチドの透過における細胞間透過経路の重要性、2)サイトカインによるTJのバリア機能調節とその機構、3)アミノ酸輸送に及ぼすサイトカインの影響の3点について検討した。 1.細胞間透過経路の重要性 食品タンパク質由来のトリペプチドにはアンジオテンシン変換酵素(ACE)阻害活性をもつものがあり、それらは経口投与でも血圧上昇抑制効果を示すと言われている。しかしながら、その小腸上皮細胞層の透過機構については不明な点が多い。そこでこのようなトリペプチドがどのような透過経路で小腸上皮細胞層を透過するかをACE阻害ペプチド(Val-Pro-Pro)を用いて検討する事にした。小腸上皮細胞層のモデルとしては、TJやプロトン依存性ペプチドトランスポーターの発現が確認されているヒト結腸ガン由来株化細胞Caco-2を用いた。管腔側よりペプチドを加え、一定時間後、基底膜側の溶液に含まれるペプチド及び遊離アミノ酸量を測定した。まず、細胞内輸送経路の重要性を検討するため、トランスサイトーシスやペプチドトランスボーターに対する阻害剤の影響を調べたところ、トリペプチドの細胞層透過量に変化は観察されず、細胞内輸送経路はインタクトなトリペプチドの細胞層透過には重要でない事が示唆された。そこでもう一つの細胞層透過経路である細胞間経路の関与について更に検討を進めた。その結果、トリペプチドの管腔側への添加濃度を上昇させると基底膜側への透過速度が直線的に増加しつづけること、基底膜側から加えた時の透過速度が、管腔側から加えた時の透過速度と変化がないことなどが見出され、トリペプチドは細胞間を拡散により透過していると推定された。このように、カルシウムなどの低分子物質ばかりでなく、トリペプチドのような分子の小腸上皮細胞層透過経路としても細胞間透過経路は重要であることが示唆された。 2.サイトカインによる細胞層透過性変化 細胞間透過経路が生理活性トリペプチドの腸管上皮細胞層透過に重要な役割を持つ事が示唆されたが、この細胞間の透過性を規定しているのはTJである。近年、腸管上皮細胞の基底膜側には様々なサイトカインの受容体が発現し、上皮細胞がサイトカインから刺激を受容することによりその機能性を変化させる事が分かってきた。そのなかでTJもサイトカインによる影響を受け、それにより細胞間の物質透過性が変化する事が報告されている。しかしながら、その調節機構については、いまだ未知の部分が多い。そこで、サイトカインによるTJの調節の機構解明を試みた。 まず、サイトカインとしてインターフェロン (IFN)、インターロイキン1 (IFN)、インターロイキン1 、腫瘍壊死因子 、腫瘍壊死因子 をCaco-2細胞層の基底膜側よりそれぞれ添加し刺激した。その後細胞間透過性の指標として経上皮電気抵抗値(TER)を測定したところ、IFNで細胞層を刺激することにより有意にTERが低下することが分かった。IFNによるTER低下が実際に細胞間透過性を増加させているかを確認するため、前章で用いたトリペプチドの透過量を測定した。その結果、Caco-2細胞層をIFNで刺激すると、無刺激時と比較して著しくトリペプチドの透過性が増大した。このようにIFNは腸管上皮細胞層の透過性を亢進しうる事が示唆されたので、次にIFNにょるCaco-2細胞層透過性調節の機構を探ることにした。 をCaco-2細胞層の基底膜側よりそれぞれ添加し刺激した。その後細胞間透過性の指標として経上皮電気抵抗値(TER)を測定したところ、IFNで細胞層を刺激することにより有意にTERが低下することが分かった。IFNによるTER低下が実際に細胞間透過性を増加させているかを確認するため、前章で用いたトリペプチドの透過量を測定した。その結果、Caco-2細胞層をIFNで刺激すると、無刺激時と比較して著しくトリペプチドの透過性が増大した。このようにIFNは腸管上皮細胞層の透過性を亢進しうる事が示唆されたので、次にIFNにょるCaco-2細胞層透過性調節の機構を探ることにした。 IFNにより刺激された腸管上皮細胞は誘導型一酸化窒素合成酵素(iNOS)を発現することが報告されている。また、一酸化窒素(NO)の供与体であるニトロプルシドナトリウムは細胞間透過性を上昇させるという報告がある。このことからIFNによるTJの調節にはNOの関与が予想された。しかし、iNOS誘導の阻害、NO合成の阻害によってもTERの回復は観察されず、また新規NO供与体添加によってもTERの低下は観察されなかった。このことからIFN刺激によるTJ調節には他の経路が関与していることが示唆された。各種阻害剤を用いて更に研究を進めた結果、IFNによるTER低下は、IFN刺激の細胞内シグナル伝達分子であるJAK2を阻害するAG490、チロシンキナーゼの阻害剤であるgenistein、タンパク質合成阻害剤のcycloheximide、カルシニューリンの阻害剤であるcyclosporin A(CsA)で抑制されることが明らかになった。CsAによる抑制効果については更に検討を加えた結果、IFNの刺激によりTJ構成タンパク質であるZO-1が細胞膜画分から消失する現象をCsAが抑制する事が分かった、。さらにIFNによるZO-1の膜画分からの消失はZO-1の発現が抑制されたためではなく、ZO-1の分解もしくは膜画分からの離脱が関与している事、この現象をCsAが抑制する事が見出された。以上の結果から、IFNによるTJ調節機構にはカルシニューリンが重要な役割を担っていることが初めて明らかになった。 3.サイトカインによるアミノ酸吸収調節機構 小腸上皮細胞層には様々なアミノ酸輸送担体が存在しており、これら輸送担体は輸送するアミノ酸の性質や輸送のための駆動力の違いにより16種類の輸送系に分類れている。アミノ酸輸送は様々な要素により制御・調節を受ける事が分かってきているが、その中の1つの制御因子としてサイトカインがある。サイトカインによるアミノ酸輸送担体制御の研究は免疫系細胞に対しては詳細に行われているが、腸管上皮細胞への影響についての情報はほとんどない。しかし、腸管における上皮細胞と免疫系細胞の相互作用を考える上で、この事象を明らかにする事はきわめて重要であると考えられる。 そこで前章で取り上げたIFNを用い、腸管上皮細胞のアミノ酸輸送に及ぼす影響について研究を行った。透過性膜上に21日間単層培養したヒト腸管上皮細胞Caco-2の基底膜側にIFNを添加し6日間培養後、放射性ラベルした各種アミノ酸を管腔側より添加してその取り込み量を測定した。また、IFNによる有意な効果が観察されたものについては、更に輸送のKinetics、IFN処理の時間・濃度依存性や各種阻害剤の影響について検討を加えた。 その結果、IFNをCaco-2に作用させることによって、グルタミン酸の取り込み活性がコントロールに比べ有意に低下することが見出された。この変化は、IFN処理の時間・濃度に比例していた。また、Kineticsの測定からグルタミン酸取り込み活性はIFN処理によりVmax値、Km値ともに低下することが分かった。またcycloheximide、cyclosporin AによりIFNの効果が有意に抑制されることから、タンパク質合成レベルでの調節が起こっていること、この取り込み制御機構にもカルシニューリンが関与していることが示唆された。このことから免疫系細胞より分泌されるサイトカインがパラクラインに作用して腸管上皮細胞のアミノ酸輸送を制御する機構が存在することが分かった。 まとめ 腸管上皮細胞層の物質透過において細胞間透過経路の重要性が明らかにされた。また、腸管上皮細胞層が腸管免疫系細胞より放出されるIFNにより調節され、栄養素透過性・吸収能が影響を受ける可能性が示唆された。また、その調節機構の鍵となる分子の1つにカルシニューリンがあることも本研究によって初めて示された。本研究は、生体レベルで行うと排除すべき因子が多く、詳細な機構解析が困難である現象をCaco-2を用いたモデル系によって解析した1例であり、このような培養細胞系の有用性を示したものと考えている。今後、腸管の上皮細胞と免疫系の相互作用を「栄養素吸収への影響」という視点からさらに検討していきたい。 | 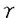 の影響
の影響