免疫系は生体内に侵入してくる外来物質を厳格に識別し、それを排除するために適切な応答を示す。すなわち免疫系は外来抗原に対して一様な応答を示すわけではなく、その抗原の性状に応じて質の異なる応答をしている。食品アレルギーを例にとると、食品抗原によってアレルギー反応の誘起のしやすさに違いがあること、アレルゲンによって異なる臨床症状が認められることが知られている。また感作量の違いによって症状が軽減することも(減感作)、悪化することもある。このような現象は抗原の構造及び量的な違いが免疫応答に深く関与していることを示している。したがってこれを解明することは抗原特異的な免疫応答の制御を考えるうえで非常に重要である。 抗原特異的な免疫応答において中心的な役割を果たしているのがT細胞である。T細胞は細胞表面上に発現するT細胞抗原レセプター(TCR)を介して抗原ペプチドと主要組織適合遺伝子複合体(MHC)分子を認識すると、TCRやその他の細胞表面分子を介してT細胞内にシグナルが伝達され、T細胞は増殖・分化し、サイトカイン産生、B細胞の活性化誘導など様々な機能を発揮する。このようにT細胞の活性化にはTCR/ペプチド/MHC分子の3分子複合体の形成が不可欠である。近年、T細胞応答はこのTCR/ペプチド/MHC分子複合体相互作用のわずかな違いにより質的に大きく変化することが報告されている。すなわち抗原ペプチドの構造や濃度のわずかな違いにより質的に異なるT細胞応答が誘導される。しかしこれまでの研究は、既に抗原に感作されたT細胞の応答を観察しているものが主であり、未だ特異的抗原に遭遇していない抗原未感作T細胞が初めて刺激を受けた時の応答(1次応答)が抗原の質的・量的な違いによりどのように変化するかについては明らかにされていなかった。 そこで本研究では卵アレルゲンである卵白アルブミン(OVA)の323-339残基領域(OVA323-339)をMHCクラスII分子であるI-Ad拘束的に認識するTCRを発現するトランスジェニックマウス(Tgマウス)を利用した。このTgマウスのT細胞の大部分はOVA323-339特異的な単一のTCRを発現しているため、生体内で抗原に感作させることなく、in vitroでの1次応答、さらにはその後の機能分化を解析することが可能となる。本研究ではこのTgマウス由来の抗原未感作CD4+T細胞の1次応答が抗原ペプチドの構造や量の違いによりどのように変化するか、またT細胞応答変化の分子機構についての解析を行った。 第1章抗原未感作CD4+T細胞の抗原ペプチドの1アミノ酸置換アナログに対する1次応答の解析 抗原未感作CD4+T細胞は特異的な抗原により活性化されると、異なる機能を持った二つのタイプの細胞へと分化する。すなわちインターロイキン(IL)-2、インターフェロン(IFN)-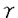 などを産生し、細胞性免疫を担うTh1型細胞と、IL-4、IL-5などを産生し、液性免疫に重要なTh2型細胞である。抗原未感作CD4+T細胞からTh1/Th2型細胞への機能分化は1次刺激時の抗原構造や抗原量に大きな影響を受けることが知られている。しかし抗原未感作CD4+T細胞が初めての抗原刺激に対してこのようなサイトカイン産生パターンを示しうるのか、またサイトカイン産生以外にどのような活性化パターンを示すのかについては明らかにされていなかった。 などを産生し、細胞性免疫を担うTh1型細胞と、IL-4、IL-5などを産生し、液性免疫に重要なTh2型細胞である。抗原未感作CD4+T細胞からTh1/Th2型細胞への機能分化は1次刺激時の抗原構造や抗原量に大きな影響を受けることが知られている。しかし抗原未感作CD4+T細胞が初めての抗原刺激に対してこのようなサイトカイン産生パターンを示しうるのか、またサイトカイン産生以外にどのような活性化パターンを示すのかについては明らかにされていなかった。 そこでTCRとの結合に関与していると考えられるOVA323-339上のアミノ酸残基を別のアミノ酸に置換したアナログペプチドを計40種類用意し、各アナログペプチド(5 M)に対する抗原未感作CD4+T細胞の1次応答、すなわち増殖応答、サイトカイン産生、in vitro抗体産生誘導能を検討した。その結果、抗原未感作CD4+T細胞のアナログペプチドに対する1次応答は次の4つのタイプに分類することができた。すなわち、(A)強いTh1型のサイトカイン産生と強い増殖応答を示すが、抗体産生は誘導しない、(B)強いTh2型のサイトカイン産生と強い増殖応答を示すが、抗体産生誘導は弱い、(C)弱いTh2型のサイトカイン産生と弱い増殖応答を示し、強い抗体産生を誘導する、そして(D)サイトカイン産生は誘導せず、増殖応答も弱いが、強い抗体産生を誘導する、という4つのタイプである。 M)に対する抗原未感作CD4+T細胞の1次応答、すなわち増殖応答、サイトカイン産生、in vitro抗体産生誘導能を検討した。その結果、抗原未感作CD4+T細胞のアナログペプチドに対する1次応答は次の4つのタイプに分類することができた。すなわち、(A)強いTh1型のサイトカイン産生と強い増殖応答を示すが、抗体産生は誘導しない、(B)強いTh2型のサイトカイン産生と強い増殖応答を示すが、抗体産生誘導は弱い、(C)弱いTh2型のサイトカイン産生と弱い増殖応答を示し、強い抗体産生を誘導する、そして(D)サイトカイン産生は誘導せず、増殖応答も弱いが、強い抗体産生を誘導する、という4つのタイプである。 これより抗原未感作CD4+T細胞は初めて抗原刺激を受けてすぐに明確なTh1/Th2型のサイトカイン産生パターンを示しうることが明らかとなった。また抗原未感作CD4+T細胞は明確なサイトカイン産生を示さなくても抗体産生誘導という活性化パターンを示すことも明らかとなった。 第2章抗原構造・量の違いが抗原未感作CD4+ T細胞の1次応答及び機能分化に及ぼす影響の解析 次にOVA323-339及びその1アミノ酸置換アナログの濃度の違いが抗原未感作CD4+T細胞の1次応答に及ぼす影響について検討した。OVA323-339は濃度によって異なるT細胞応答を誘起した。すなわち低濃度(0.005M)ではサイトカイン産生は検出されなかったが、強いin vitro抗体産生誘導が観察された。一方中濃度(0.05M)ではTh2型のサイトカイン産生が、高濃度(0.5M以上)ではTh1型のサイトカイン産生が誘導された。低濃度刺激で観察された抗体産生誘導は濃度の上昇に伴い低下した。 第1章でB、C、Dタイプの応答を誘導した1アミノ酸置換アナログを用いた場合にも、OVA323-339と同様に濃度変化に伴うT細胞応答の変化が認められた。すなわち濃度上昇に伴い、抗体産生誘導、Th2型サイトカイン産生というT細胞応答の変化が観察された。これらのアナログペプチドはOVA323-339と同程度の増殖応答を誘導するためには100倍から1000倍の濃度が必要とされたことから、アミノ酸置換によりTCRとの親和性が大きく低下したことが予想された。 以上より抗原未感作CD4+T細胞の1次応答は抗原ペプチド/MHC分子複合体とTCRとの親和性に加え、抗原ペプチド/MHC分子複合体の密度によって決定されることが示唆された。この二つの要素を掛け合わせたものはavidityと称されているが、低avidityでは抗体産生誘導が、中avidityではTh2型サイトカイン産生が、高avidityではTh1型サイトカイン産生が誘導されるものと考えられた。 さらにこのような抗原未感作CD4+T細胞の1次応答変化がTh1/Th2型細胞への分化に与える影響を解析したところ、低濃度、中濃度のOVA323-339及び高濃度のアナログペプチドはTh2型細胞への分化を、高濃度のOVA323-339はTh1型細胞への分化を誘導した。したがって、抗原未感作CD4+T細胞の1次応答の質の違いがその後のTh1/Th2細胞への機能分化を決定することが判明した。 第3章抗原濃度の違いにより生じる抗原未感作CD4+ T細胞応答変化の機構解析 次に抗原濃度の違いにより生じる抗原未感作CD4+T細胞の1次応答変化の機構解析をOVA323-339を抗原として用いて行った。細胞表面分子の発現はフローサイトメトリーあるいは定量RT-PCR法で、サイトカインmRNA及びサイトカイン遺伝子発現に関与する転写因子mRNAの発現量は定量RT-PCR法で調べた。 低濃度刺激では細胞表面分子に関してはT細胞活性化マーカーであるCD69、CD25の発現に加え、B細胞活性化誘導分子であるOX40、CD40リガンド(CD40L)の発現が観察された。またIL-4 mRNA及びTh2型サイトカイン遺伝子の発現に重要な転写因子であるGATA-3、c-maf mRNAの発現も誘導された。抗IL-4、抗CD40L、抗OX40抗体の添加により抗体産生誘導が強く抑制されたことから、これらの分子が低濃度のOVA323-339が誘導する抗体産生応答に重要な役割を果たしていることが明らかとなった。 中濃度刺激では細胞表面分子の発現はさらに強くなり、IL-4、GATA-3、c-maf mRNAの高発現が認められた。また低濃度刺激では観察されなかったFasリガンド(FasL)の発現も観察された。一方、高濃度刺激ではOX40、CD40L、FasLに加え、IFN-の強い発現が誘導されたが、GATA-3、c-maf、IL-4 mRNAの発現は認められなかった。中・高濃度刺激ではB細胞にFas-FasL系を介したアポトーシスが誘導されることが判明し、また抗FasL及び抗IFN-抗体の添加により抗体産生応答が誘導された。したがって、中・高濃度刺激ではT細胞が産生するIFN-と、Fas-FasL系を介して誘導されるB細胞のアポトーシスがin vitro抗体産生応答に負に働いていることが判明した。 これより抗原未感作CD4+T細胞の細胞表面分子、サイトカイン発現に関与する転写因子の発現量は抗原濃度によって異なることが判明し、これがT細胞の機能的な応答の変化を生み出す原因であることが示唆された。 以上の研究により、抗原未感作T細胞は抗原構造や濃度のわずかな違いに応じて異なる1次応答を示すことが明らかとなった。これより生体に侵入してくる抗原の質的・量的な違いは、生体内免疫応答の質的な相違を誘起し、さらにはアレルギーなど免疫疾患の誘起のしやすさにも大きな影響を及ぼすことが示唆された。本研究で得られた知見は抗原特異的なT細胞の活性化機構の解明、さらには抗原特異的な免疫抑制法の解明に大きく寄与するものと思われる。 |