ゲノムから遺伝情報を取り出す転写は遺伝子発現制御の最も重要な段階である。真正細菌のRNAポリメラーゼは、コア酵素とこれにプロモータ選択能及び転写開始能を付与するシグマ( )因子とからなる。本研究は、大腸菌の定常期・浸透圧特異的なシグマ因子、38に特異的な機能領域を解析した結果をまとめたものであり、4章よりなる。 )因子とからなる。本研究は、大腸菌の定常期・浸透圧特異的なシグマ因子、38に特異的な機能領域を解析した結果をまとめたものであり、4章よりなる。 序章で大腸菌を中心として得られている真正細菌のRNAポリメラーゼ、シグマ因子、プロモータ等についてのこれまでの知見を総括した後、第1章では、大腸菌で比較的最近発見された定常期並びに浸透圧応答性の主要型シグマ因子,38(rpoS遺伝子産物)の特異的な機能領域を検索した結果について述べている。38蛋白質は主要因子70と共通する基本骨格構造を持つが、領域4.2よりC-末端に至る領域(CTE:C-terminal element)は両者の間では相同性が低い。ところが、さまざまな真正細菌のrpoS相同遺伝子産物間では保存性が高いことから、それぞれ特有の機能を持つことが示唆された。このことを検証するため、野生型38とCTE欠失型38(1-314)による細胞内転写活性への影響をkatE、lac及びlacUV5プロモーターを用いて調べた。その結果、38(1-314)は、in vitroで転写開始活性を保持しているにも拘わらず、in vivoでkatE、fic、wrbAなどの38特異的なプロモーターの転写を行なうことができないことが分かった。一方、38は定常期における遺伝子発現の正の制御因子であると同時に70依存型プロモーターからの転写開始を抑制する働きを持つ(負の制御)ことが知られている。実際、rpoS欠損株ではいくつかの遺伝子発現の定常期における抑制が解除される。そこで、この負の制御に38-CTEが関与しているかどうかについて70と38の両方によって認識されるlacプロモーターを用いて調べた。その結果、38(1-314)を持つ株では、lacプロモーターからの定常期における転写抑制が解除されていることが示された。これらの結果は、38のCTE領域がin vivoにおける38による正と負の制御に関与していることを示唆している。更に、16アミノ酸からなるCTE領域のどのアミノ酸がこれらの機能に関与しているかをアラニンスキャンニング法により解析した。その結果、317番目のロイシン、323番目のアスパラギン、327番目のロイシンの比較的良く保存された3つのアミノ酸が重要であることが明らかになった。 第2章では、38のCTE領域が38とコア酵素との結合能に影響している可能性について検討を行った結果を述べている。それぞれ精製したコア酵素と野生型並びにCTE欠失型の38蛋白質とを1:1,1:4,1:16の量比で混合し、形成されるホロ酵素と転写能を解析した。その結果、CTE欠失38の量比を増やすと転写活性能が回復することが明らかになった。CTEがホロ酵素の安定性に寄与をしていることを示唆している。 第3章は、38のCTE領域の有無とin vitroにおける転写の塩濃度感受性について解析した結果を述べている。野生型のE38ではK-glutamate(あるいはNa-glutamate)濃度が100-200mMと上昇するにしたがってficプロモーターからの転写が促進されるのに対し、E38(1-314)では50mM K-glutamateが最適濃度で、200mMでは転写が著しく阻害された。尚、それぞれの条件で形成されるホロ酵素の量には顕著な差異は認められなかった。これらの結果は38のCTE領域が細胞内の塩濃度に対応した活性制御に関わっていることを示唆しており、定常期細胞では細胞内塩濃度が100-500mMであるとする知見と一致する。 第4章は総合討論である。 以上要するに本論文は、大腸菌の定常期特異的シグマ因子38固有の機能を持つCTE領域を見い出し、この領域が塩濃度感受性と主要シグマ因子70に対する負の制御に関わっていることを見い出したものであり、学術上、応用上寄与するとことが少なくない。よって審査委員一同は、本論文が博士(農学)の学位論文として価値あるものと認めた。 | 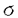 38の機能領域解析
38の機能領域解析