糖尿病性腎症は糖尿病三大合併症の一つであり、1998年にはわが国で透析導入の必要な腎不全の原因の第一位を占めている。しかし、その病態の詳細なメカニズムは解明されておらず、適切なモデル動物の開発が必要である。 APAハムスターは6ヶ月齢頃より軽度の腎糸球体硬化症を自然発症する。また、この動物はストレプトゾトシン(SZ)の単回投与により長期にわたり持続する高血糖および高脂血症を呈するため、糖尿病合併症の動物モデルとして高く評価されている。このモデルにおける糖尿病性腎症の糸球体病変の発生機構を解明するため、本論文では病態の変化を追うと同時にその様々な増悪因子について検索した。 第I章・第1節(I-1)では基礎データとしてAPAハムスターの自然発症腎病変を組織学的に解析した。第I章・第2節(I-2)ではSZ誘発糖尿病APAハムスターの血液、尿の臨床病態と、腎病変に関する病理組織学的検索を行った、第II章では腎糸球体病変の発症機構を明らかにするため、第1節(II-1)では糸球体病変の発症に関わる因子について組織化学的・免疫組織化学的手法を用いて検索し、第2節(II-2)で病変形成に関わると思われるサイトカインや成長因子の発現の変化をRT-PCR法を用いて調べた。 I-1)APAハムスターの腎糸球体ではメサンギウム領域(MR)の軽度の拡張と糸球体基底膜(GBM)の肥厚が加齢に伴い進行していた。この病変にはメサンギウム細胞が産生する細胞外基質とmesangial interpositionが深く関わっていることが免疫組織化学的検索および電顕検索により示唆された。 I-2)SZ投与後24時間目から血糖値および血中トリグリセリド値が、48時間目から血中コレステロール値が上昇し、実験期間中(SZ投与後6ヶ月)持続した。糖尿病群では経時的な腎機能障害の進行が認められた。病理組織学的には糖尿病群で腎糸球体の肥大に引き続きMRの拡張やGBMの肥厚といった、ヒトと類似の病変がみられた。また、泡沫細胞も観察された。 II-1)組織化学的、免疫組織学的検索の結果、以下のことが明らかになった。1)糸球体内の細胞数の増加。2)この増加は糸球体内の細胞増殖とマクロファージの浸潤によるものであることが示唆された。泡沫細胞の一部はマクロファージであることが示された。3)性状の変化したメサンギウム細胞に発現する -smooth muscle actin(SMA)陽性細胞数は対照群と比して糖尿病群で多かった。また、泡沫細胞の一部もSMA陽性を示した。4)糖尿病群では全体にレクチン染色性が向上し、最終糖化産物も沈着していた。5)高脂血症の発症とともに糸球体や尿細管間質への脂質の沈着が始まり、経時的に増えていた。泡沫細胞内および大小の動脈壁にも脂質の沈着が観察された。脂質の沈着部位には免疫染色によりapoEおよびBの蓄積が観察された。6)投与後1、3および6ヶ月目にはIV型コラーゲン、ファイプロネクチンおよびラミニンの増加が病変部でみられた。さらに、糖尿病群ではI型およびIII型コラーゲンの顕著な蓄積が観察された。 -smooth muscle actin(SMA)陽性細胞数は対照群と比して糖尿病群で多かった。また、泡沫細胞の一部もSMA陽性を示した。4)糖尿病群では全体にレクチン染色性が向上し、最終糖化産物も沈着していた。5)高脂血症の発症とともに糸球体や尿細管間質への脂質の沈着が始まり、経時的に増えていた。泡沫細胞内および大小の動脈壁にも脂質の沈着が観察された。脂質の沈着部位には免疫染色によりapoEおよびBの蓄積が観察された。6)投与後1、3および6ヶ月目にはIV型コラーゲン、ファイプロネクチンおよびラミニンの増加が病変部でみられた。さらに、糖尿病群ではI型およびIII型コラーゲンの顕著な蓄積が観察された。 II-2)RT-PCR法を用いて病変形成に関連すると考えられる各種サイトカイン・増殖因子のmRNAの発現について検索した。急性期の腎皮質ではIL-6、IL-1および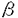 mRNAの増加が観察された。1および3ヶ月日にはIL-6、PDGF BおよびTGFのmRNA発現が増加していた。 mRNAの増加が観察された。1および3ヶ月日にはIL-6、PDGF BおよびTGFのmRNA発現が増加していた。 以上の結果からSZ誘発糖尿病APAハムスターの糖尿病性腎症は(1)高血糖、脂質、AGEによる糸球体構成細胞の代謝障害や、糸球体の肥大、過濾過などの血行動態の異常により、メサンギウム細胞の増殖やこの細胞の性状に変化が生じる。(2)腎構成細胞や浸潤してきたマクロファージなどにより産生されるサイトカインがメサンギウム細胞による細胞外基質などの産生を促す。(3)細胞外基質の沈着および脂質やAGEを含んだ泡沫細胞により糸球体構造が異常を来たし、高度な蛋白尿や腎機能の低下がおこる、というメカニズムで生じていると考えられた。このようにして形成された胃病変はヒトの糖尿病性腎症の瀰漫型病変と非常に類似していた。 以上、本論文はAPAハムスターにおける糖尿病性腎症の臨床病態、腎糸球体病変の特徴およびその発症メカニズムを明らかにしたものであり、APAハムスターがヒトの糖尿病性腎症の有用なモデル動物であることを示した。また、その発症メカニズムの詳細な経時的解析により、いくつかの新たな知見が得られた。これらの知見は今後の糖尿病性腎症研究に大いに貢献するものである。よって審査委員一同は、本論文が博士(獣医学)の学位論文として価値あるものと認めた。 |