| 内容要旨 | | 神経細胞は高度な極性を持った細胞であり,たくさんの樹状突起と1本の長い軸索を持っている。中には細胞体からシナプス端までの長さが1mを越えるものもある。ほとんどの蛋白質は細胞体中で合成されるので,軸索やシナプスで使われる物質は特別に分化した輸送機構なしには効率よく運搬できない。神経細胞には軸索輸送という特殊な細胞内輸送機構が発達しており,これは速度により速い流れ(数百ミリ/日)と遅い流れ(数ミリ/日)に分けられる。このうち前者は主に小胞や膜小器官の輸送に,後者は細胞骨格蛋白,蛋白複合体などの運搬にあずかっているとされている。一般に軸索輸送は細胞骨格蛋白のひとつである微小管を軌道とする大別して2種類のモーター蛋白により行われている。順行性の輸送にはキネシンスーパーファミリー(KIFs)の大部分の分子が,逆行性の輸送にはダイニンスーパーファミリーの一群が関係していることが今までの研究により明らかにされてきた。今までの研究では主として軸索輸送にあずかっている分子それ自身の性質(形態学的,生化学的,生物物理的)の解明がなされてきたが,それぞれの分子の生理学的・細胞生物学的意味は不明であった。近年,遺伝子組換え法などによりKIFs分子の機能,換言すればどのような分子あるいは小胞を運んでいるかという事が明らかにされつつある。本研究はマウスの標的遺伝子組換え法を用い,最も古典的なKIFsのひとつであるKIF1Bが細胞内でどのように機能しているかという問題についての研究である。 KIF1Bはマウス脳cDNAから単離されたKIFのひとつで,多くのKIFsがとる二量体構造ではなく単量体ではたらくモーター蛋白質であるKIF1サブファミリーに属し,微小管依存性ATPase活性を担うモータードメインをそのN末端側に持っている。 さらに最近になって,KIF1Bはオルタナティプスプライシングによってモータードメインのみを共有する異なった2種類のモーター,KIF1B-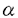 (分子量121kD)とKIF1B- (分子量121kD)とKIF1B-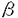 (分子量199kD)からなることがわかってきた。KIF1B-はKIF1Aに非常によく似ている。KIF1Aは軸索内でシナプス小胞の前駆体の輸送を行うという報告がある。一方KIF1Bについては,神経細胞内においてミトコンドリアの局在と一致する,グルコーストランスポーター1結合蛋白(GLUT1-CBP)と結合する,という報告がある。しかしこれらの仕事は培養細胞や酵母の系でなされたものであり,実際の生体細胞内での機能,それが個体レベルでどのように反映されるかという点についてはわかっていなかった。そこで今回私は標的遺伝子組換え法を用いてkif1B遺伝子を欠失させたマウスを解析し,KIF1B分子の機能の一部を解明した。この際用いたターゲティングベクターでは,モーター蛋白質の必須エレメントであるATP結合部位を欠失させる設計がとられているため,上述のKIF1B一とKIF1B-の双方を一度に欠失させることができた。従って表現型も両者の機能の損失を反映したものとなった。 (分子量199kD)からなることがわかってきた。KIF1B-はKIF1Aに非常によく似ている。KIF1Aは軸索内でシナプス小胞の前駆体の輸送を行うという報告がある。一方KIF1Bについては,神経細胞内においてミトコンドリアの局在と一致する,グルコーストランスポーター1結合蛋白(GLUT1-CBP)と結合する,という報告がある。しかしこれらの仕事は培養細胞や酵母の系でなされたものであり,実際の生体細胞内での機能,それが個体レベルでどのように反映されるかという点についてはわかっていなかった。そこで今回私は標的遺伝子組換え法を用いてkif1B遺伝子を欠失させたマウスを解析し,KIF1B分子の機能の一部を解明した。この際用いたターゲティングベクターでは,モーター蛋白質の必須エレメントであるATP結合部位を欠失させる設計がとられているため,上述のKIF1B一とKIF1B-の双方を一度に欠失させることができた。従って表現型も両者の機能の損失を反映したものとなった。 具体的には,マウスkif1B遺伝子cDNAのATP結合コンセンサスであるP-loop周辺を含む断片をプローブとして2個の独立なgenomicクローンを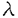 ファージライブラリから単離した。これらのクローンを用い,P-loopを含むエクソンとその周辺領域をpoly(A)を含まないIRESLacZpPGKneoカセットで置き換える形で,ポジティブネガティブ選別法によるターゲティングベクターを構築した。これは,poly(A)トラップ法により相同組換え体の選択効率の改善を図ると共に,kif1B遺伝子の発現箇所を同定できるようにlacZ遺伝子を組み込んだものである。このベクターをJ1系のES細胞に電気穿孔法を用いて導入し,G418により選別したクローンについてSouthern blotting法によるスクリーニングを行い,630個のG418耐性コロニーから13個の相同組換え体を得た。次にこのうち4個をマウス胚盤胞に注入することで独立なES細胞クローンに由来する2系統のマウスを得,ヘテロ接合体同士をかけ合わせることによってkif1B遺伝子欠損マウスを得た。なお,表現型は2系統間で同じであることを確認した。 ファージライブラリから単離した。これらのクローンを用い,P-loopを含むエクソンとその周辺領域をpoly(A)を含まないIRESLacZpPGKneoカセットで置き換える形で,ポジティブネガティブ選別法によるターゲティングベクターを構築した。これは,poly(A)トラップ法により相同組換え体の選択効率の改善を図ると共に,kif1B遺伝子の発現箇所を同定できるようにlacZ遺伝子を組み込んだものである。このベクターをJ1系のES細胞に電気穿孔法を用いて導入し,G418により選別したクローンについてSouthern blotting法によるスクリーニングを行い,630個のG418耐性コロニーから13個の相同組換え体を得た。次にこのうち4個をマウス胚盤胞に注入することで独立なES細胞クローンに由来する2系統のマウスを得,ヘテロ接合体同士をかけ合わせることによってkif1B遺伝子欠損マウスを得た。なお,表現型は2系統間で同じであることを確認した。 ヘテロ接合体は野生型にくらべて外見・行動ともに変化なかった。そのかけ合わせの結果ほぼメンデル比の割合でホモ接合個体が産まれたが,これらはいずれも体が小さく(体重約20%減)筋緊張の低下,刺激に対する応答の低下を呈した。またチアノーゼを示し,出生後30分以内で死亡した。解剖して肺を水中に入れると,野生型・ヘテロ接合体のものは中に気泡がみられ水に浮いたが,ホモ接合体のものは気泡がなく沈んだままだった。光学顕微鏡で観察すると肺胞の拡張がみられず,新生仔は自発呼吸をしていないことが明らかとなった。 自発呼吸が起きない理由を調べるため,出産直前にあたる18.5日胚を用い呼吸に関係する部位のうち(1)筋・骨格系,(2)神経筋接合部,(3)下位呼吸運動ニューロン(脊髄前角),(4)上位呼吸中枢(脳幹)のどこが冒されているかを検討した。まず肺・肋間筋・横隔膜を光学顕微鏡・電子顕微鏡で観察したが,明らかな異常は認められなかった。横隔膜の神経筋接合部の電気生理学的解析の結果も正常個体との間に顕著な差はなかった。一方,脊髄前角については横隔神経の神経細胞体があるC4レベルを光顕・電顕て観察したところ,ともに顕著な変化を認めた。ホモ接合体では野生型に比して単位面積当たりのシナプスの数およびシナプス内のシナプス小胞の数ともに統計的に有意な減少がみられた。脳幹についても明らかな変化が認められ,呼吸に関係した神経核である延髄背側網状核・延髄腹側網状核・孤束核・腹外側網状核・舌下神経核において,kif1B欠失個体ではいずれも大きさ・ニューロンの数ともに減少していた。もっともこの傾向は呼吸関連に限定されたものではなく,顔面神経核にもやはり同様の減少が見られた。個々のニューロンについては,ニッスル小体の染色が弱く,神経変性の徴候と考えられた。 次に,中枢神経系の異常を細胞レベルで調べるため,海馬の神経細胞培養を行った。KIF1B欠失ニューロンは正常なニューロンよりはるかに生存率が悪く,培養開始から5日後には90%が死滅した。さらに神経突起の伸長が遅く,正常細胞よりステージの発達が明らかに遅れていた。 これらの結果から,kif1B遺伝子産物は,神経細胞の分化および維持に重要な働きを果たしており,またその結果,ノックアウトマウスでは呼吸中枢を含む中枢神経系の多くの神経核群の発達が極度に遅延し中枢性の自発呼吸の欠如が起こったことが示唆される。このマウスは新生児無呼吸の一つのモデルとなることが期待される。 さらに,海馬神経細胞・上衣細胞・胚体外膜細胞の初代培養を使ってミトコンドリアの分布を調べた。しかしKIF1Bの有無による違いは特に認められなかった。これは,もうひとつのミトコンドリアのモーターとして知られるKIF5Bによって補償されていたことが考えられる。またGLUT1-CBPとKIF1Bの結合について,免疫細胞化学によってGLUT1の分布を調べたが,特に分布に変化はみられなかった。 総体として,kif1Bノックアウトマウスの培養細胞における表現型はkif1Aのそれと類似していた。この結果はKIF1B-とkif1Aが高い相同性を持っているということによって説明される。KIF1Aはシナプス小胞前駆体を輸送していることが明らかになっているが,KIF1B-についてもこれと類似の機能が示唆され,KIF1B-が神経細胞の形作り,機能発現および生存にとって基本的役割を果たしていることが示された。 |