本研究は高等真核生物の染色体DNA複製開始制御において重要な役割を果たしていると考えられるASK(Activator of S phase Kinase)タンパク質の機能を明らかにするために、ASK遺伝子のプロモーター領域を単離し、ASK遺伝子の転写制御機構の解析を行ったものであり,下記の結果を得ている。 1.ヒト正常線維芽細胞株であるWI-38細胞、マウスインターロイキン3(muIL-3)依存性に増殖するpro-B cell株であるBa/F3において、ASK遺伝子の発現が増殖刺激によって誘導され、S期に進行していくにつれて徐々に上昇していくことがノーザンブロッティングにより示された。 2.ヒトT細胞株であるJurkat細胞のゲノムDNAを種々の制限酵素で切断し、サザンブロッティングによりASK遺伝子を解析した結果、弱いバンドが多数検出され、ASK遺伝子には疑遺伝子や類似遺伝子が存在することが示唆された。 3.ASK遺伝子の5’領域、約5kbのDNA断片を単離し、ASK遺伝子の5’領域に結合しうる転写因子の結合配列を検索したところ、5個のE2F結合モチーフと6個のSp1結合モチーフが密集する領域がみつかった。この領域を含む約1.5kbのDNA断片をルシフェラーゼ遺伝子の上流につないだレポータープラスミドを構築し、HeLa、NIH3T3、Ba/F3細胞に一過性にトランスフェクションしたところ、ASK遺伝子の5’領域を含まないルシフェラーゼ遺伝子のみのものをトランスフェクションしたときに比べ、極めて強いルシフェラーゼ活性を示した。また、このレポータープラスミドをNIH3T3細胞に一過性にトランスフェクションし、血清飢餓状態(G0期)から血清刺激により細胞周期を進行させたとき、ルシフェラーゼ活性が徐々に上昇し、これが内在性ASK遺伝子の増殖刺激による変化とほぼ一致すること示した。これらの結果より、ASK遺伝子の上流約1.5kbのDNA断片はASK遺伝子の発現を制御するプロモーター領域であることが示された。 4.Ba/F3細胞にレポータープラスミドとE2F1,2,3いずれかを発現するベクターを一過性に共発現させ、ルシフェラーゼ活性を測定したところ、E2F発現ベクターを共発現させると共発現しないものと比べ、極めて高いルシフェラーゼ活性を示した。また、E2Fの結合すると考えられる配列を持っていないレポータープラスミドもE2Fによって活性化された。この結果はこれまでに同定されたE2F結合配列以外の新規のE2F結合配列の存在、E2Fによる未知の転写活性化機構(他の転写因子等を介した)の存在を示唆する。さらに、E2F結合配列に変異を導入したレポータープラスミドでは血清刺激、外来性E2Fによるプロモーター活性の上昇が抑えられた。 5.E2F1タンパク質を発現する組み換えアデノウイルスを静止期のラット線維芽細胞株REF52に感染させ、E2F1により内在性のASK mRNAが誘導されることをノーザンブロッティングで示した。 6.N端にFlagタグを融合させたASKタンパク質を恒常的に発現するBa/F3細胞を樹立し、この細胞株を増殖因子(muIL3)非存在下で培養した時、親株より静止期の状態に入りにくい(S期の細胞の割合が高い)ことを示した。このことは、一時的ではあるがASKタンパク質の高発現が増殖刺激なしに細胞をS期に進行させることができることを示唆している。 以上,本論文は高等真核細胞のDNA複製開始制御に重要な役割を果たしていると考えられるASKの転写制御にはE2F転写因子が関与しており、ASKタンパク質の過剰発現が増殖刺激なしに細胞をS期に進行させることを示した。これまでにE2Fの強制発現により静止期の線維芽細胞をS期に進行させることができ、このS期進行にはE2Fの転写活性化作用が必須であることはわかっていたが、E2FによるS期誘導の分子機構については、数多くのE2F標的遺伝子が同定されているにもかかわらず、どの標的遺伝子がS期誘導の中心的役割を果たしているかは不明であった。本研究はASKタンパク質がE2FによるS期誘導の中心的役割を果たしている可能性を示唆しており,高等真核生物のDNA複製開始制御機構の解明に重要な貢献をなすものであり、学位の授与に値するものと考えられる。 | 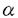 、thymidine kinase(TK)、cyclinE、E2F1、Orc1、Cdc6など)の遺伝子発現を制御していることがわかっている。また、E2Fの強制発現により静止期の線維芽細胞をS期に進行させることができ、このS期進行にはE2Fの転写活性化作用が必須である。しかし、E2FによるS期誘導の分子機構については、これまでに数多くのE2F標的遺伝子が同定されているにもかかわらず、どの標的遺伝子がS期誘導の中心的役割を果たしているかは、はっきりしていない。ASKタンパク質は先に述べたようにS期の開始、進行に重要な役割を果たしていると考えられることから、ASK遺伝子の発現がE2Fによって制御され、E2FによるS期誘導の中心的役割を果たしている可能性が高いと考えられる。
、thymidine kinase(TK)、cyclinE、E2F1、Orc1、Cdc6など)の遺伝子発現を制御していることがわかっている。また、E2Fの強制発現により静止期の線維芽細胞をS期に進行させることができ、このS期進行にはE2Fの転写活性化作用が必須である。しかし、E2FによるS期誘導の分子機構については、これまでに数多くのE2F標的遺伝子が同定されているにもかかわらず、どの標的遺伝子がS期誘導の中心的役割を果たしているかは、はっきりしていない。ASKタンパク質は先に述べたようにS期の開始、進行に重要な役割を果たしていると考えられることから、ASK遺伝子の発現がE2Fによって制御され、E2FによるS期誘導の中心的役割を果たしている可能性が高いと考えられる。