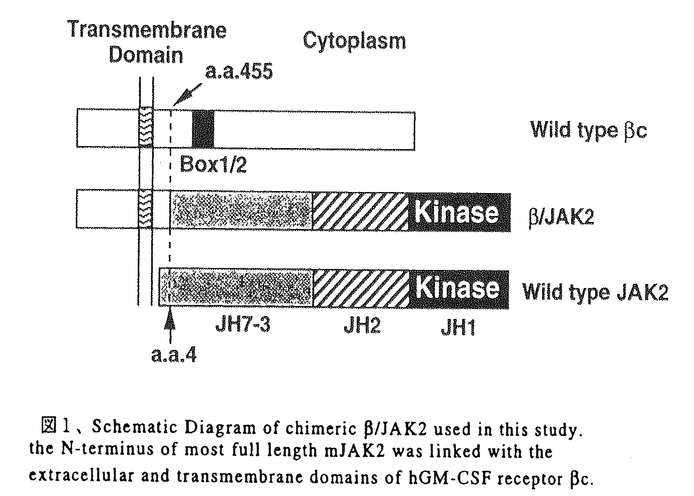
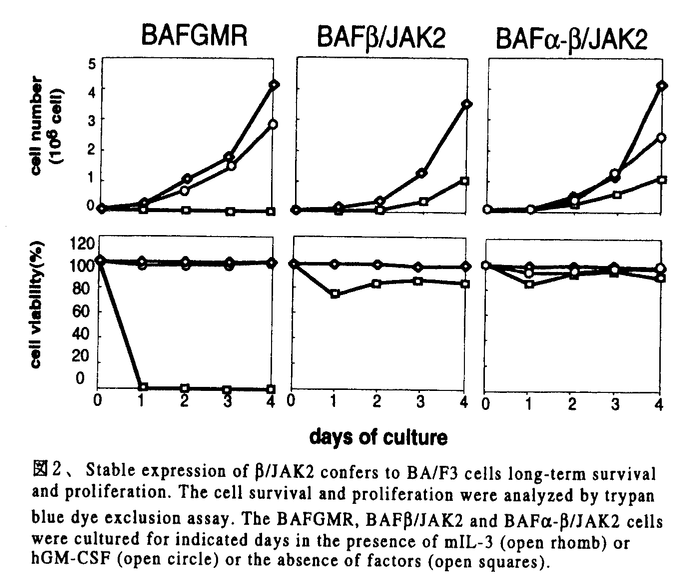
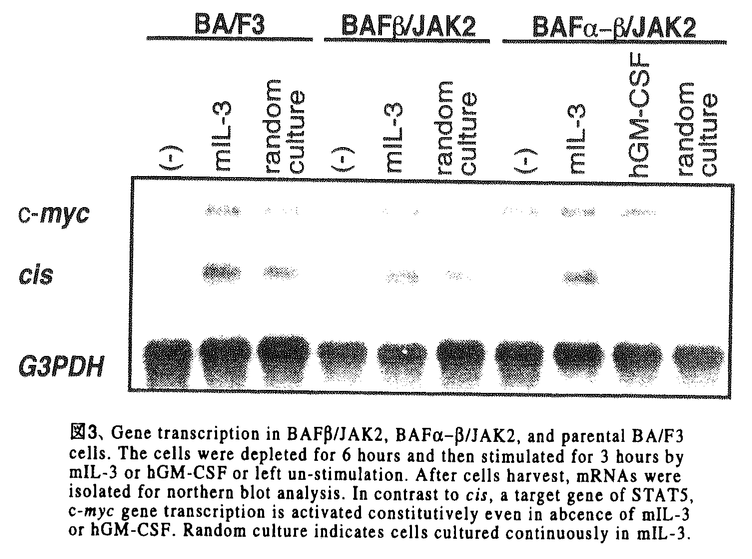
本研究は血液系細胞の生存、増殖を制御するサイトカインシグナル伝達におけるJAK2の役割の解明を目的として、サイトカインレセプターの細胞内領域の関与なしにJAK2のみの活性を解析できる系、すなわちhGM-CSFレセプター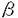 鎖の細胞外と膜貫通領域にを結合させたキメラ分子/JAK2を構築して、細胞内で発現させ、このキメラ分子による細胞機能制御とシグナル伝達能を検討し、以下の結果を得ている。 鎖の細胞外と膜貫通領域にを結合させたキメラ分子/JAK2を構築して、細胞内で発現させ、このキメラ分子による細胞機能制御とシグナル伝達能を検討し、以下の結果を得ている。 1)BA/F3細胞に/JAK2を単独で発現させた細胞株BA/F/JAK2,および/JAK2をhGM-CSFレセプター 鎖と共に発現させた細胞株BA/F-/JAK2を樹立した。これらの細胞内での/JAK2の活性化状態をチロシンリン酸化を指標に解析したところ、いずれの細胞においても/JAK2はhGM-CSF、mIL-3刺激の有無にかかわらず、構成的にチロシンリン酸化されていた。これまでに鎖がhGM-CSFの非存在下でも二量体を形成しており、この構成的な鎖二量体の形成には鎖の細胞外、膜貫通領域で十分であることが明らかとなっていることから、/JAK2キメラ分子は鎖部分を介した二量体形成によりJAK2部分が活性化されていると予測された。このことはJAK2が二量体化により活性化されるというモデルともよく一致するものであった。 鎖と共に発現させた細胞株BA/F-/JAK2を樹立した。これらの細胞内での/JAK2の活性化状態をチロシンリン酸化を指標に解析したところ、いずれの細胞においても/JAK2はhGM-CSF、mIL-3刺激の有無にかかわらず、構成的にチロシンリン酸化されていた。これまでに鎖がhGM-CSFの非存在下でも二量体を形成しており、この構成的な鎖二量体の形成には鎖の細胞外、膜貫通領域で十分であることが明らかとなっていることから、/JAK2キメラ分子は鎖部分を介した二量体形成によりJAK2部分が活性化されていると予測された。このことはJAK2が二量体化により活性化されるというモデルともよく一致するものであった。 2)BAF/JAK2およびBAF-/JAK2細胞株の増殖能について、トリパンブルー色素を用いた細胞数計測により検討したところ、両細胞株ともにmIL-3非依存性に生存し、かつ細胞数も長期的に増加した。また/JAK2キメラ分子はmIL-3枯渇条件下でのゲノムDNAの断片化を抑制し、一方3Hチミジンの取り込みを誘導した。以上の結果から、鎖細胞内ドメインの関与なしにJAK2の活性化のみでBA/F3細胞の生存、増殖誘導に十分であることが明らかになった。次に/JAK2の発現による細胞内の遺伝子転写誘導を調べたところ、c-myc遺伝子の転写の構成的な増強が観察された。これに対して、c-fos、bcl-XL、cisなどの遺伝子転写の活性化は認められなかった。このことからc-myc遺伝子の転写は細胞の増殖、生存に関与していることが強く示唆された。 3)シグナル伝達分子の活性化について検討したところ、STAT5、Shc、SHP-2およびERK2のリン酸化は観察されず、これらの分子の下流で誘導される-casein、c-fosプロモーターの活性化も見られなかった。さらに、細胞の生存のシグナルへの関与が報告されているPI3キナーゼ、Aktの活性化も見い出されなかった。これらの結果から、/JAK2はSTAT5、RAS/RAF/MAPキナーゼあるいはPI3キナーゼ/Aktシグナルカスケードとは独立したシグナル経路を介して細胞の生存、増殖を誘導すると考えられた。 4)さらに、サイトカイン非存在下で生存、増殖しているBAF/JAK2、BAF-/JAK2細胞に対する種々のキナーゼ阻害剤の効果を検討した。チロシンキナーゼの阻害剤であるGenisteinはこれらの細胞の生存、増殖を抑制したが、PI3キナーゼ阻害剤のWortmanninは細胞の生存、増殖に影響を及ぼさなかった。したがって、PI3キナーゼ経路は/JAK2からの細胞の生存と増殖のシグナルに必須ではないことがさらに確認できた。Genisteinはチロシンキナーゼ阻害剤であるが、JAK2の活性に対しては影響を及ぼさないことが知られており、したがって/JAK2の下流において他のチロシンキナーゼが関与していることが強く示唆された。 以上のことからhGM-CSFレセプターとの融合によるJAK2の構成的活性化はBA/F3細胞の生存、増殖の誘導、c-mycの活性化に十分であること、鎖細胞内領域やSTAT5、MAPKカスケードの活性化はこれらの活性には必要ではないことが明らかになった。本研究により、/JAK2の系が細胞の生存、増殖に至る未知のシグナル伝達経路を探る格好の系であることが明らかになり、今後の解析によりシグナル伝達経路の詳細が解明されると考えられる。また、このキメラ分子の遺伝子導入により血液細胞の運命を改変することが可能になれば、血液系疾患のメカニズムの解明や、治療法の開発へも応用できると期待される。従って本論文は学位の授与に値するものと判断できる。 |